
BIOLOGÍA Y GEOLOGÍA




1. SISTEMAS DE COORDINACIÓN
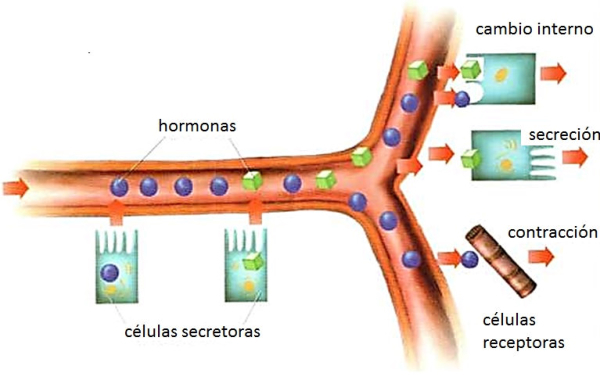
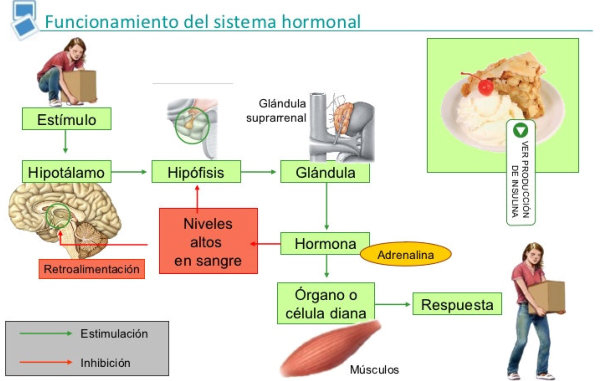
Los animales, como organismos vivos, necesitan de la función de relación, que les permite detectar cambios en el entorno y responder a ellos. para conseguirlo dispone de diferentes sistemas de coordinación. Los sistemas de coordinación se encargan de que todas las funciones corporales se realicen de forma coordinada. Son el sistema nervioso y el sistema hormonal o endocrino. Juntos forman el sistema neuroendocrino. La función de relación consiste en la captar información del medio (estímulos) y elaborar respuestas para adaptarse a los cambios del entorno. En este proceso intervienen los siguientes elementos: Estímulos: cualquier cambio en el medio que pueda ser detectado. Pueden se físicos (radiaciones, temperatura, presión) o químicos (hormonas, fármacos,…). Receptores: estructuras que captan estímulos internos o externos. Efectores: estructuras que producen respuestas (músculos, glándulas). Transmisores: llevan la información entre receptores y efectores. Pueden ser señales eléctricas o sustancias químicas (neurotransmisores, hormonas). Las respuestas de los sistemas de coordinación llevan a dos mecanismos: Comportamiento: es el conjunto de respuestas nerviosas y hormonales ante estímulos externos. En animales la repuesta más habitual es el movimiento. Homeostasis: es el conjunto de respuestas nerviosas y hormonales ante estímulos internos. La homeostasis mantiene las constantes vitales en equilibrio: presión sanguínea, temperatura, latidos, respiración, excreción, pH, etc.2. EL SISTEMA NERVIOSO
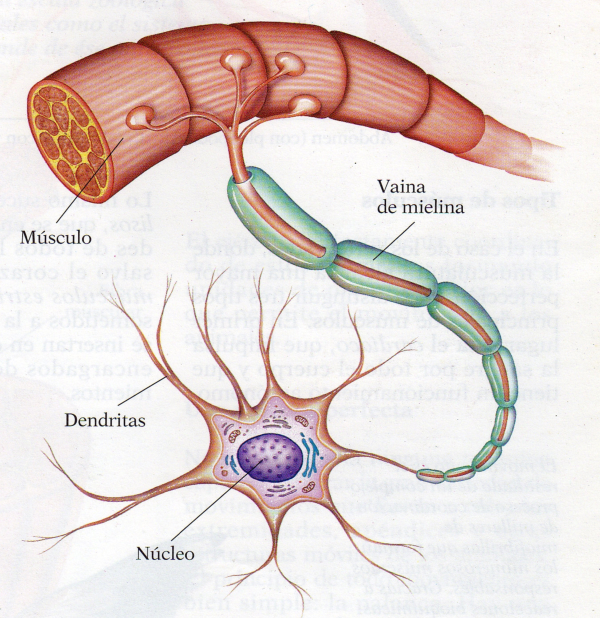
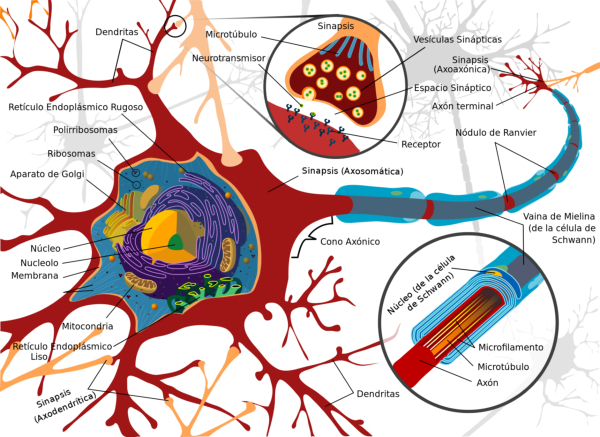
El sistema nervioso (SN) es una compleja red encargada de interpretar y almacenar la información que llega de células especializadas en captar estímulos (los receptores). Luego transmite esa información a otras células del nervioso o a los efectores, que darán respuestas (músculos y glándulas). El sistema nervioso se compone de un tejido especial, el tejido nervioso formado por dos tipos de células: neuronas y células de la glía.2.1. EXCITABILIDAD NEURONAL
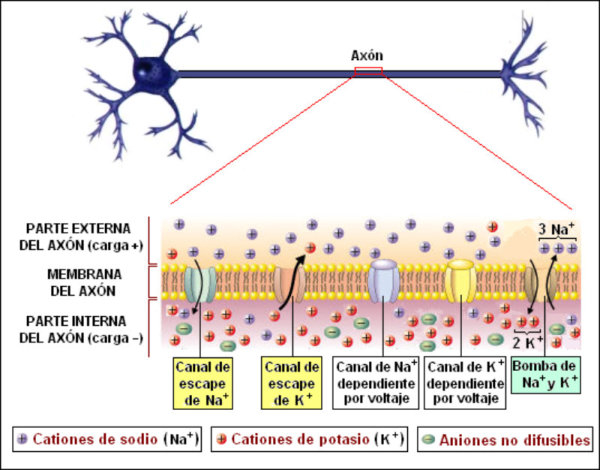
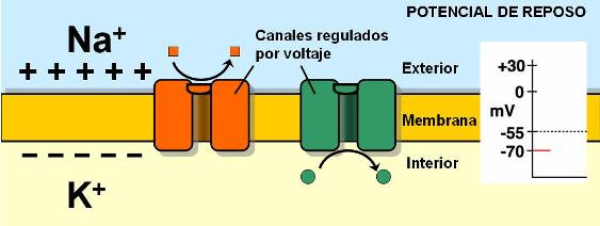
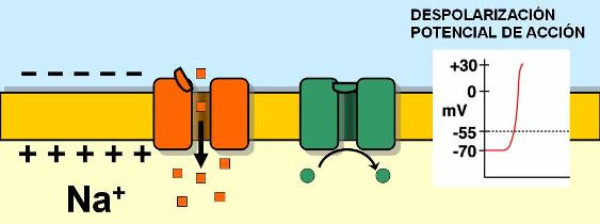
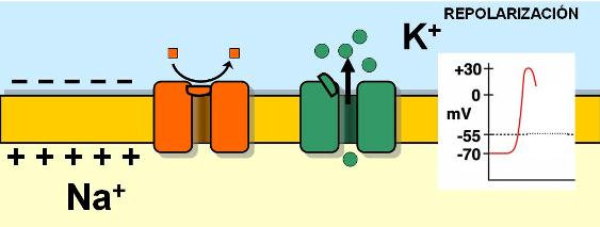
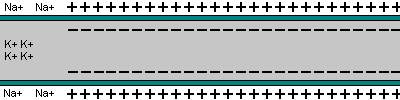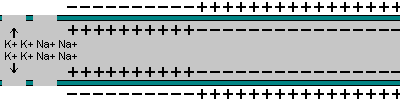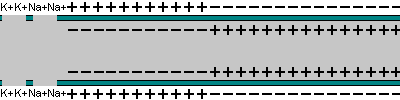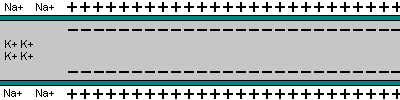
La capacidad de captar estímulos y emitir respuestas se debe a la excitabilidad neuronal, es decir, a la generación de impulsos nerviosos gracias a un potencial de membrana. 2.1.1. POTENCIAL DE MEMBRANA O DE REPOSO El potencial de membrana es la diferencia de potencial entre el exterior y el interior de la membrana plasmática. Esta diferencia se debe a una acumulación de cargas de distinto signo y es de unos -70 milivoltios (mV). El signo menos indica que en el interior hay más cargas negativas, debido al Cl - y a proteínas con carga negativa. La bomba de Na + /K + : se encuentra en la membrana plasmática y saca de la células iones Na + al tiempo que introduce iones K + , lo que provoca un desequilibrio. Canales de K + : los canales de K + se hallan siempre abiertos, por lo que el K + , muy abundantes en el interior, tienden a salir a favor de gradiente químico. Sin embargo la alta concentración de cargas negativas en el interior se opone a esta salida (gradiente eléctrico). Potencial de membrana: cuando ambos gradientes se equilibran, se alcanza el potencial en reposo o potencial de membrana, que es de unos -70 mV. 2.1.2. DESPOLARIZACIÓN: POTENCIAL DE ACCIÓN La presencia de un estímulo abre los canales de Na + y cierra los de K + . Al entrar masivamente Na + a favor de gradiente, se alcanza el umbral de excitación, que termina haciendo positivo el interior celular. Se da así una despolarización, que puede llevar el potencial a +40 mV, momento en que se alcanza el potencial de acción, que genera un impulso nervioso que se propaga por el axón hasta la siguiente neurona a través de la sinapsis. 2.1.3. REPOLARIZACIÓN La repolarización consiste en recuperar el potencial de -70 mV. Tras una excitación, se cierran los canales de Na + , se abren los de K + y vuelve a actuar la bomba de Na + /K + . Durante la repolarización (unos milisegundos) la neurona no puede reaccionar a un estímulo y se dice que está en periodo refractario.2.2. CONDUCTIVIDAD NEURONAL
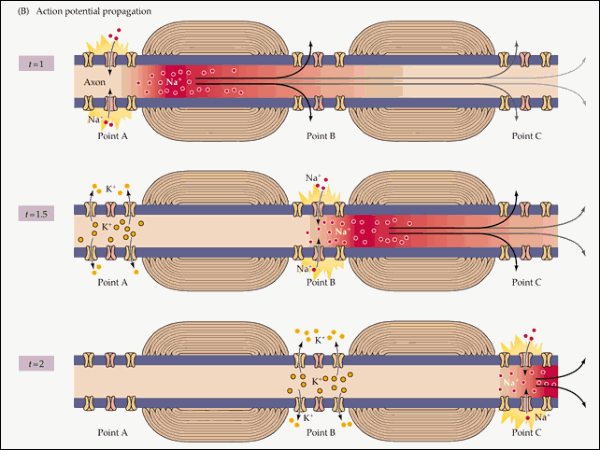
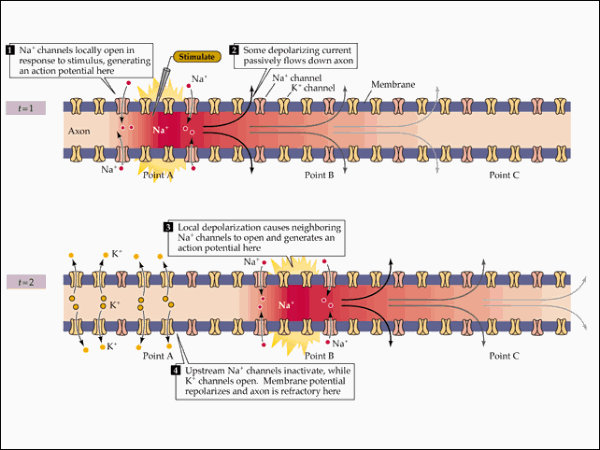
El impulso nervioso ocasionado por el potencial de acción se transmite por toda la neurona, desde las dendritas hacia el axón y los botones sinápticos. Esto se consigue con la apertura y cierre sucesivo de los diferentes canales iónicos. La despolarización se transmite de forma similar. Las neuronas más gruesas y las mielinizadas transmiten el impulso mucho más deprisa, gracias a la conducción saltatoria de un nódulo de Ranvier al siguiente.2.3. SINAPSIS
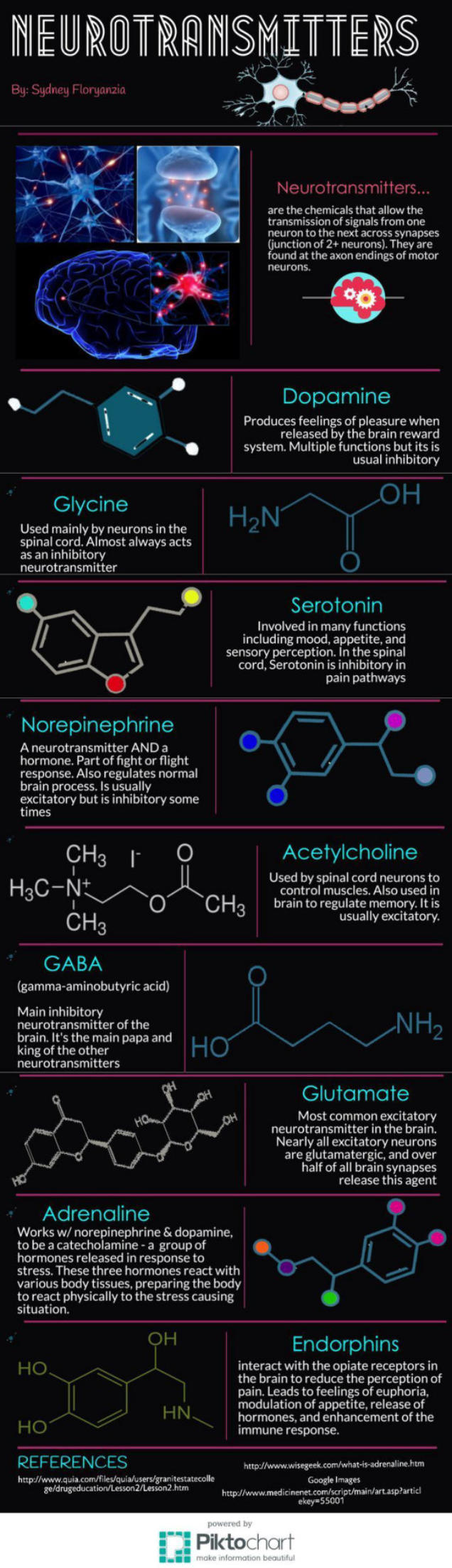
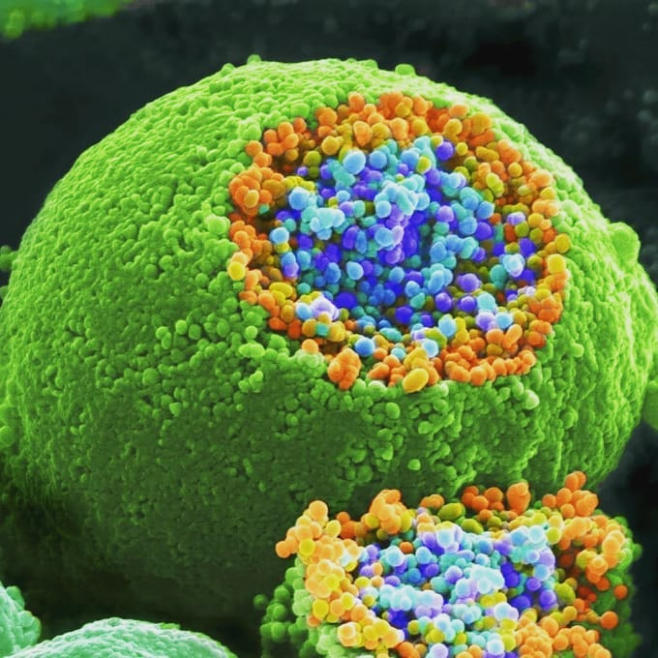
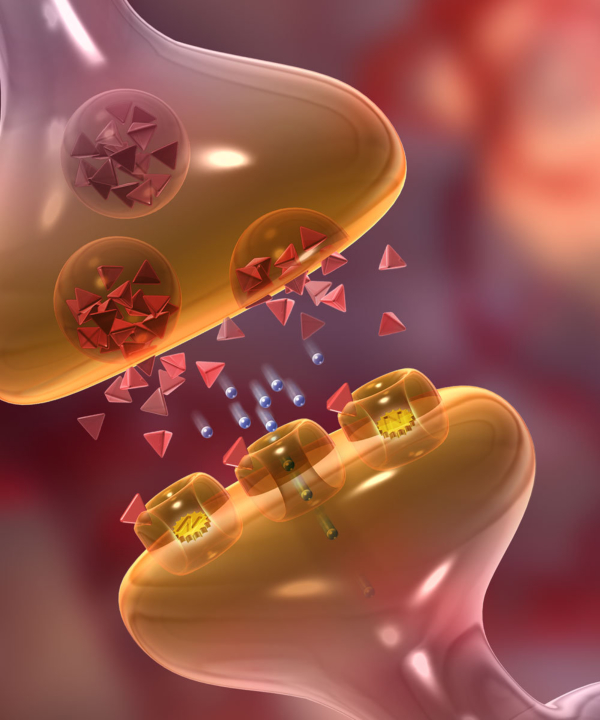
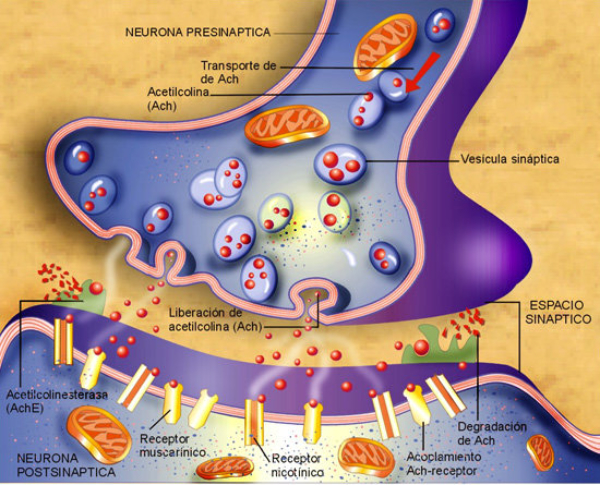
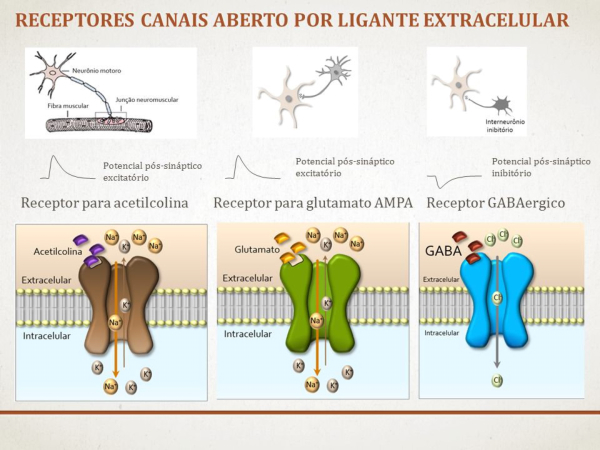
La sinapsis es la unión funcional entre dos neuronas o una neurona y un efector (músculo o glándula). Hay sinapsis eléctricas (el impulso nervioso pasa de una célula a otra libremente), pero la mayoría son químicas, el potencial de acción se transmite por sustancias químicas llamadas neurotransmisores. Éstos se sintetizan en el soma y se almacenan en vesículas sinápticas situadas en las terminaciones del axón, llamadas botones sinápticos. El potencial de acción recorre el axón de la neurona presináptica, llega a los botones sinápticos y allí se abren los canales de Ca + . El Ca + entra en los botones y provoca el vaciado de los neurotransmisores a la hendidura sináptica mediante exocitosis. Los neurotransmisores se unen a la membrana de la neurona postsináptica y provocan la apertura de sus canales iónicos: Si el neurotransmisor es excitador (noradrenalina, acetilcolina, glutamato,…) se abren los canales de Na + . El sodio entra en la neurona postsináptica, que se despolariza y da lugar a un potencial de excitación postsináptico (PEPS) que contribuye al potencial de acción. Si el neurotransmisor es inhibidor (encefalinas, endorfinas, glicina,…) se abren los canales de Cl - , que entra, o los de K + , que sale, hiperpolarizando la neurona postsináptica y genera un potencial de inhibición postsináptico (PIPS), que dificulta el potencial de acción. La neurona postsináptica realiza entonces una integración, sumando los efectos excitadores e inhibidores. Si la suma supera el umbral necesario, se genera el potencial de acción y el impulso nervioso continúa.2.4. EL SISTEMA NERVIOSO EN INVERTEBRADOS
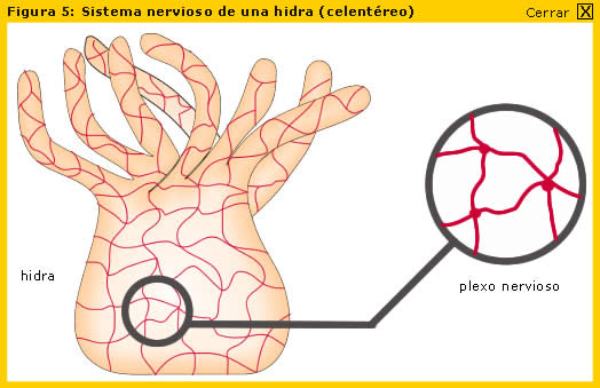
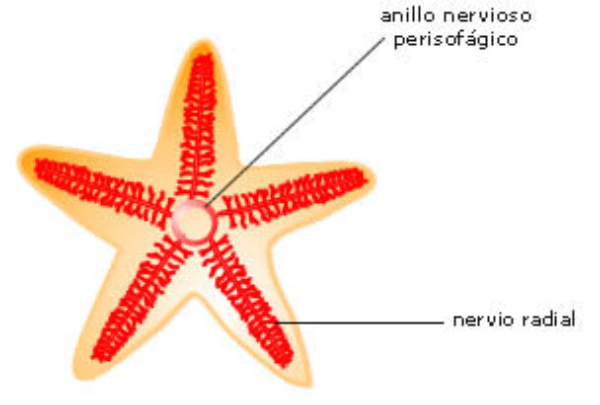
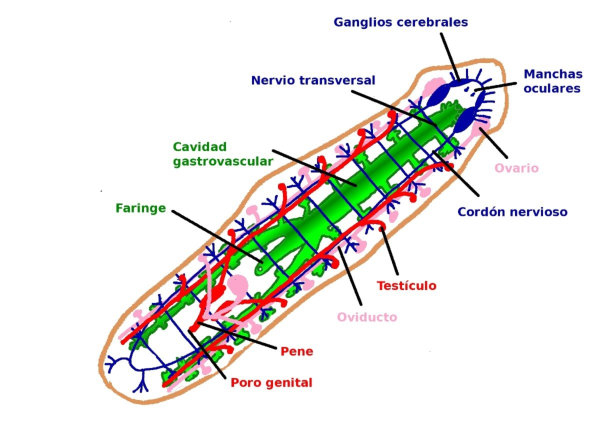
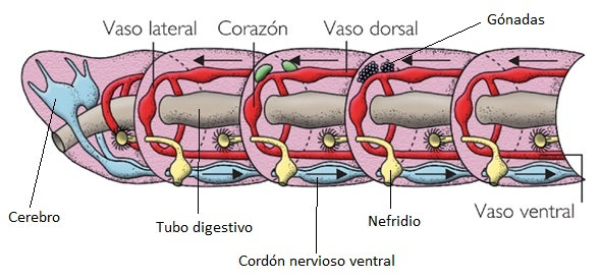
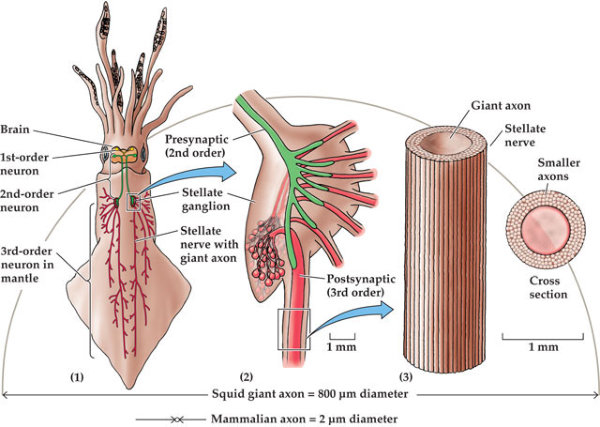
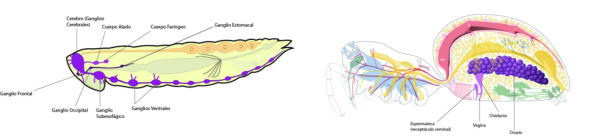
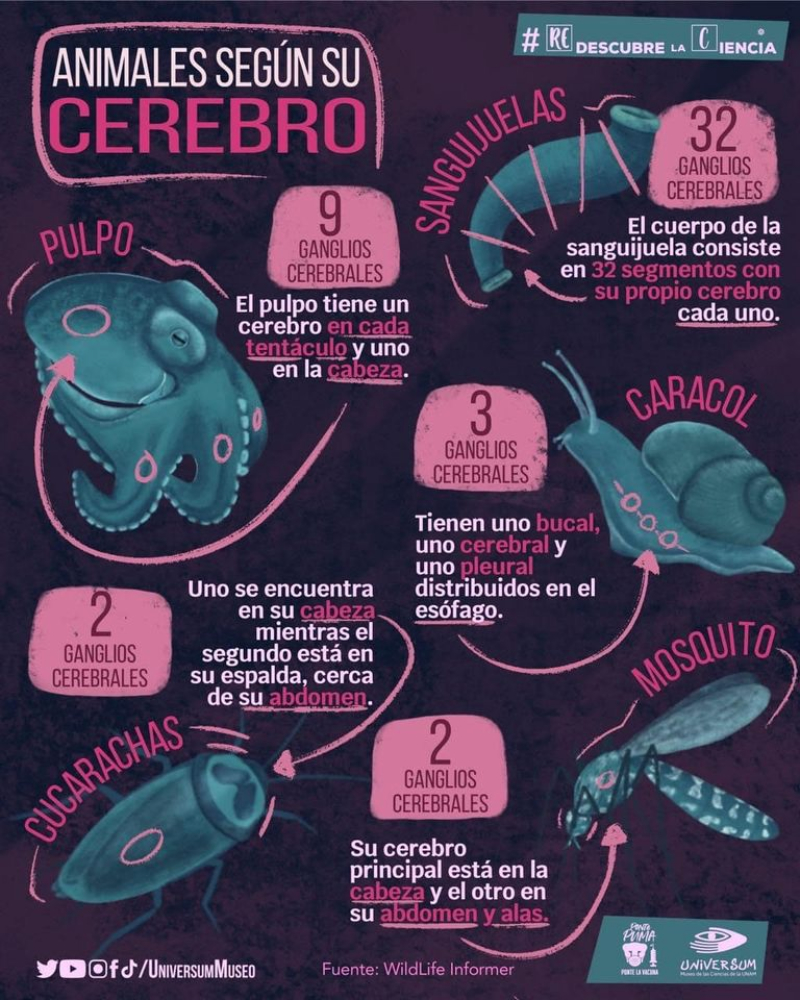
En la evolución animal se aprecia una tendencia a agrupar las neuronas en ganglios y a la cefalización, favorecida por la simetría bilateral. La cefalización llevó a la acumulación ganglionar en la parte delantera del animal, formando la cabeza, donde se acumularon los principales receptores. 2.4.1. PORÍFEROS Las esponjas no tienen un SN propiamente dicho. Sólo grupos de neuronas dispersas y sin interconexión apenas. 2.4.2. CNIDARIOS Presentan un plexo nervioso, una red neuronal sencilla y extendida por todo el cuerpo. Lo usa para la contracción celular y captura de alimento. 2.4.3. EQUINODERMOS Presentan un anillo nervioso central que rodea al esófago y del que parten cordones nerviosos radiales hacia cada brazo o región del cuerpo, originando una red nerviosa superficial. 2.4.4. PLATELMINTOS Y NEMATODOS Sistema nervioso cordal. Existen ganglios cerebrales en la cabeza de los que parten cordones nerviosos que recorren todo el cuerpo. 2.4.5. ANÉLIDOS, MOLUSCOS Y ARTRÓPODOS Sistema nervioso ganglionar. Hay ganglios cerebrales dorsales que forman un anillo alrededor del esófago (collar periesofágico). Este anillo se comunica con dos cordones nerviosos ventrales unidos por fibras transversales: cordón nervioso escaleriforme. Anélidos: un par de ganglios conectados en cada segmento. SN con aspecto de escalera. Moluscos: hay una progresión. Los más sencillos (bivalvos) tienen dos cordones con 3 a 5 pares de ganglios repartidos por todo el cuerpo. En los cefalópodos sólo hay un cordón y los ganglios se agrupan en una masa cerebral protegida por una cápsula gelatinosa. Artrópodos: la cefalización aumenta, los ganglios cerebrales forman un cerebro con varias regiones.2.5. EL SISTEMA NERVIOSO EN VERTEBRADOS
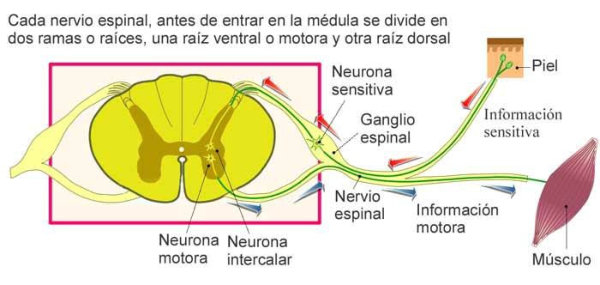
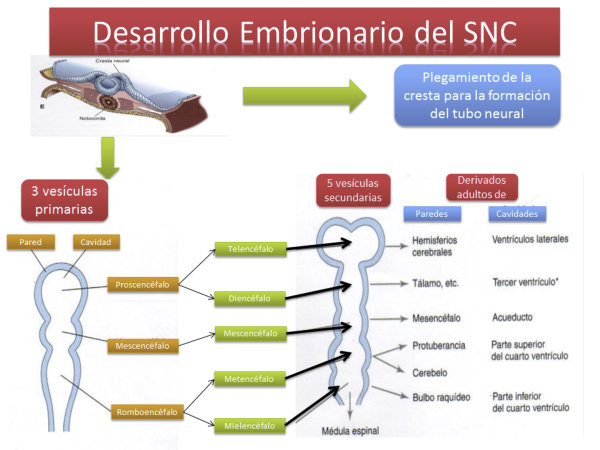
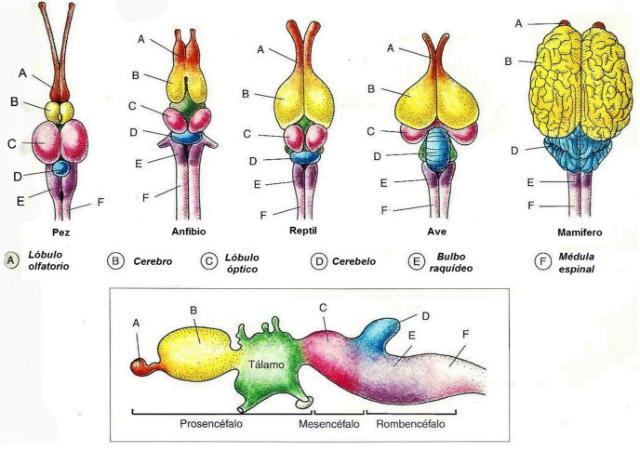
En vertebrados el procesamiento de la información se realiza principalmente en una estructura nerviosa muy compleja: el sistema nervioso central (SNC), formado por el encéfalo (encerrado en el cráneo) y la médula espinal (protegida por la columna vertebral). Del SNC parten nervios que, junto a los ganglios simples forman el sistema nervioso periférico (SNP). A partir del tubo neural del embrión se forma el cordón hueco de la médula espinal y, en su parte anterior, se ensancha para dar el encéfalo, que presenta tres regiones: Prosencéfalo: o encéfalo anterior, que incluye al cerebro. Mesencéfalo: o encéfalo medio. Rombencéfalo: que incluye el cerebelo y el bulbo raquídeo.3. RECEPTORES SENSORIALES
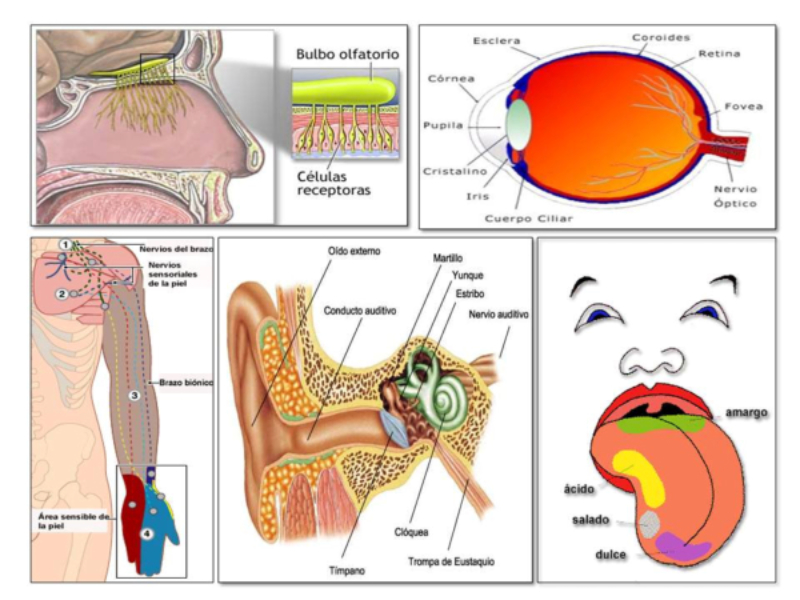
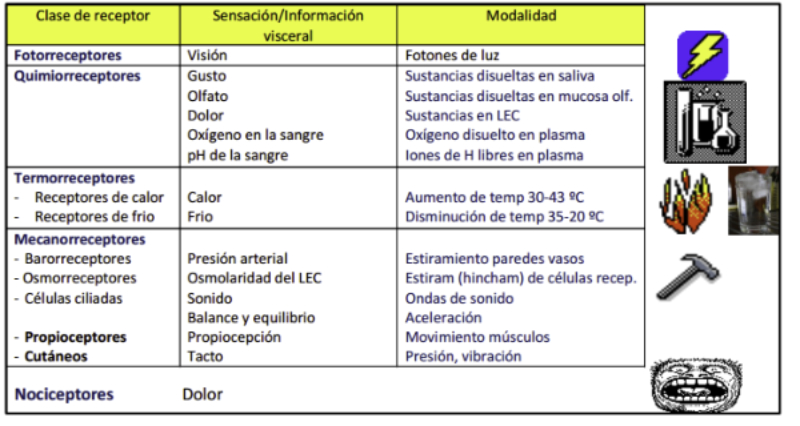
Los receptores sensoriales son células especializadas en captar estímulos. Los receptores detectan determinados tipos de energía (mecánica, térmica, química, electromagnética) y la convierten en impulsos nerviosos que las neuronas de los animales interpretan para percibir el mundo que les rodea. Dado que el número de receptores y, por tanto, de tipos de energía detectable, es limitado, los animales sólo pueden detectar una parte del mundo físico. Los receptores pueden ser exterorreceptores (captan información del exterior) e interorreceptores (captan información del interior y permiten la homeostasis). Además, según el tipo de energía que reciben los receptores se clasifican en: mecanorreceptores, termorreceptores, quimiorreceptores, magnetorreceptores, electrorreceptores y fotorreceptores.3.1. MECANORRECEPTORES
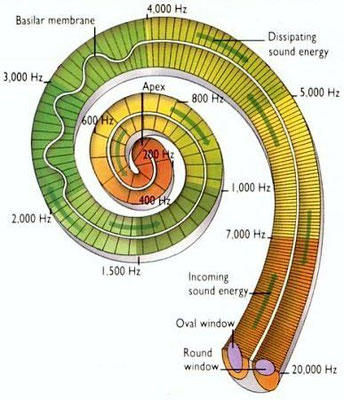
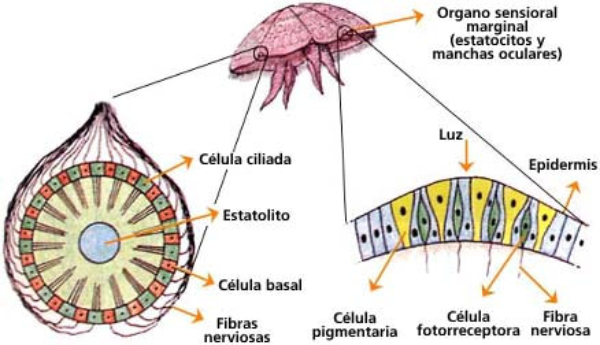
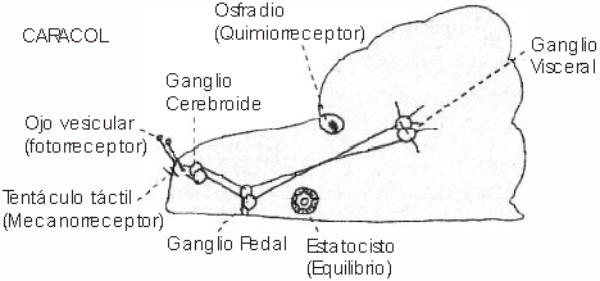
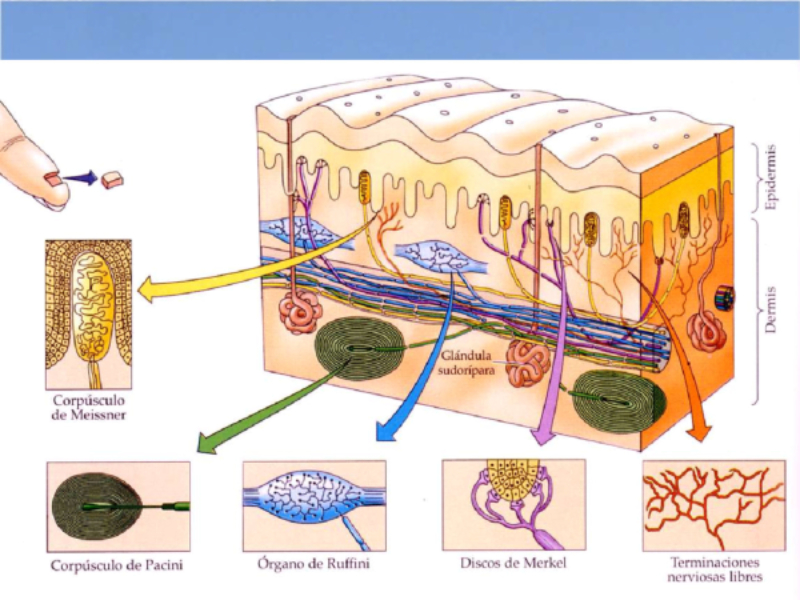
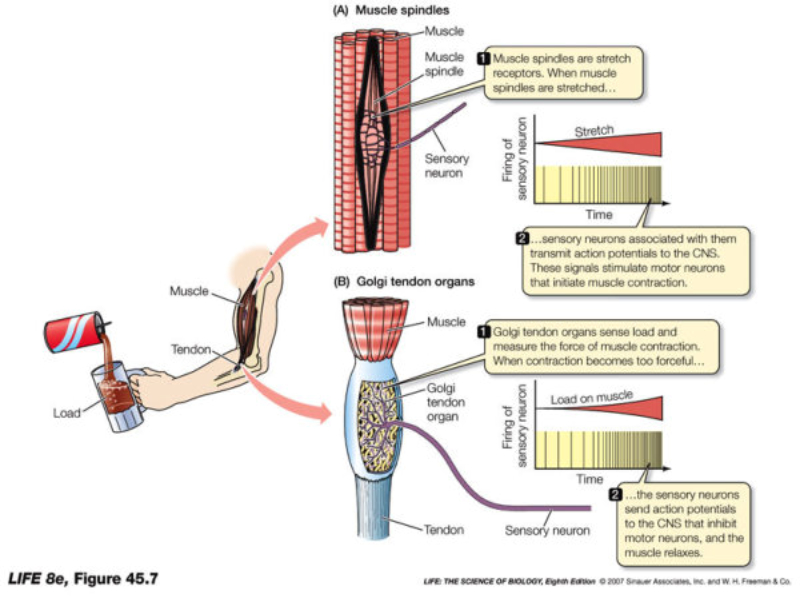
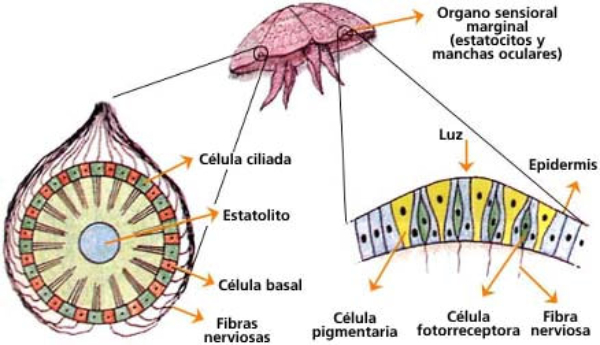
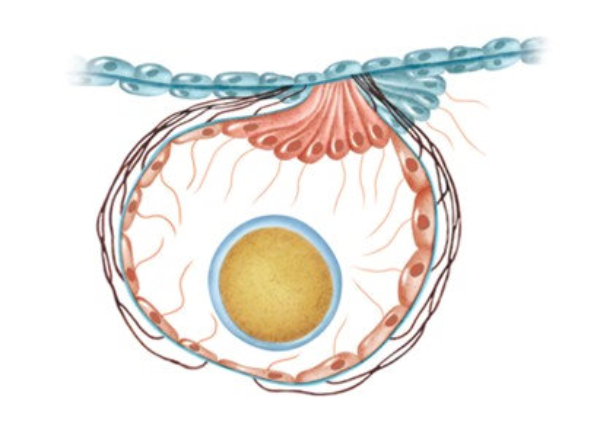
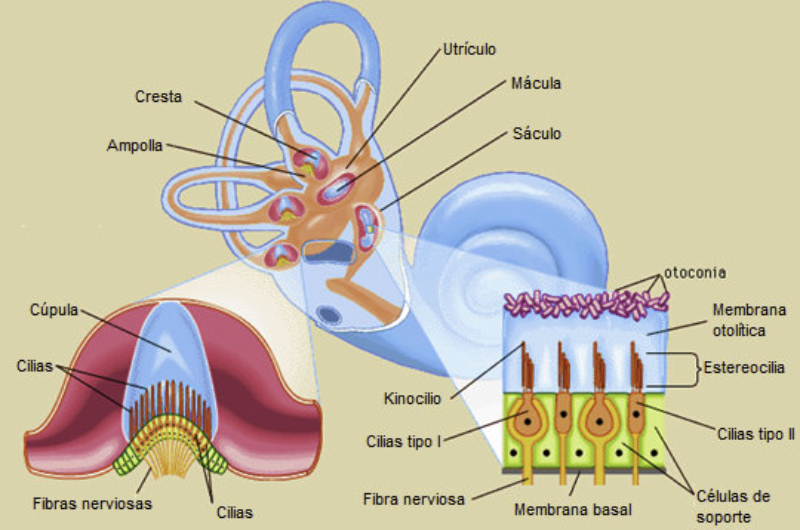
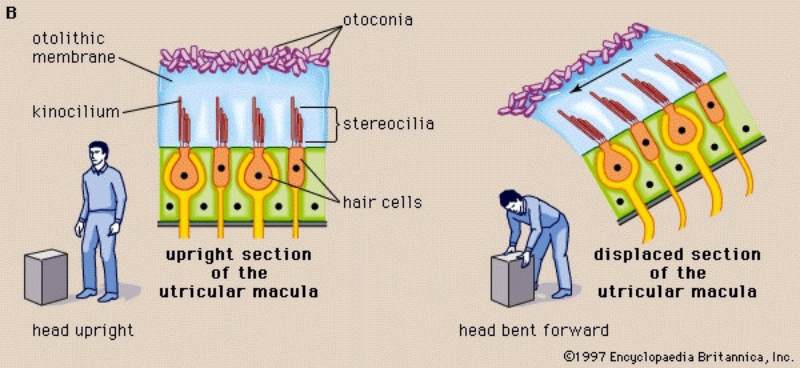
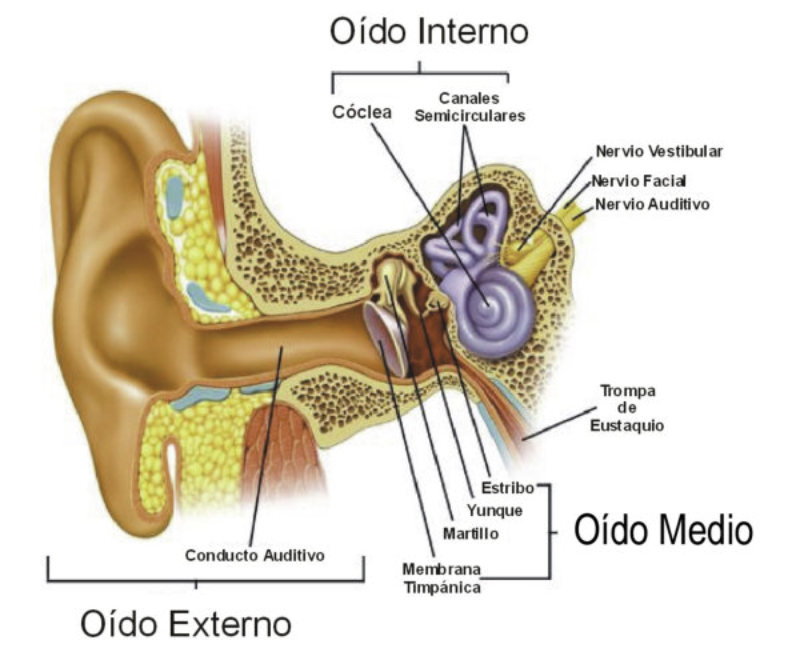
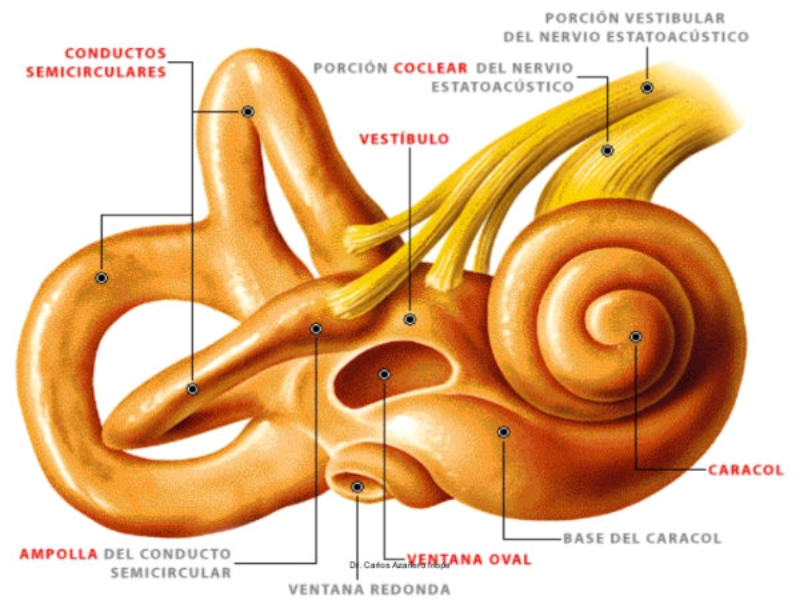
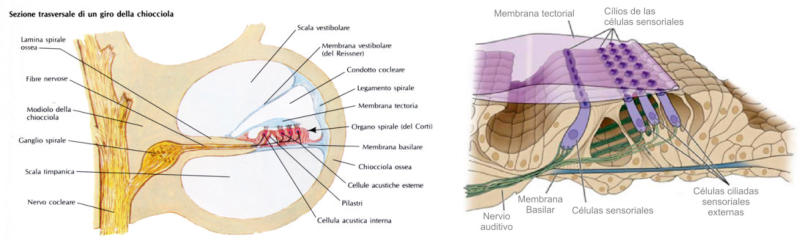
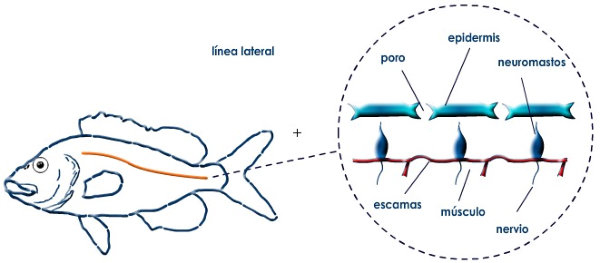
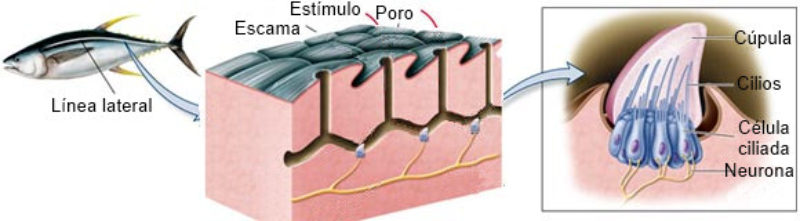
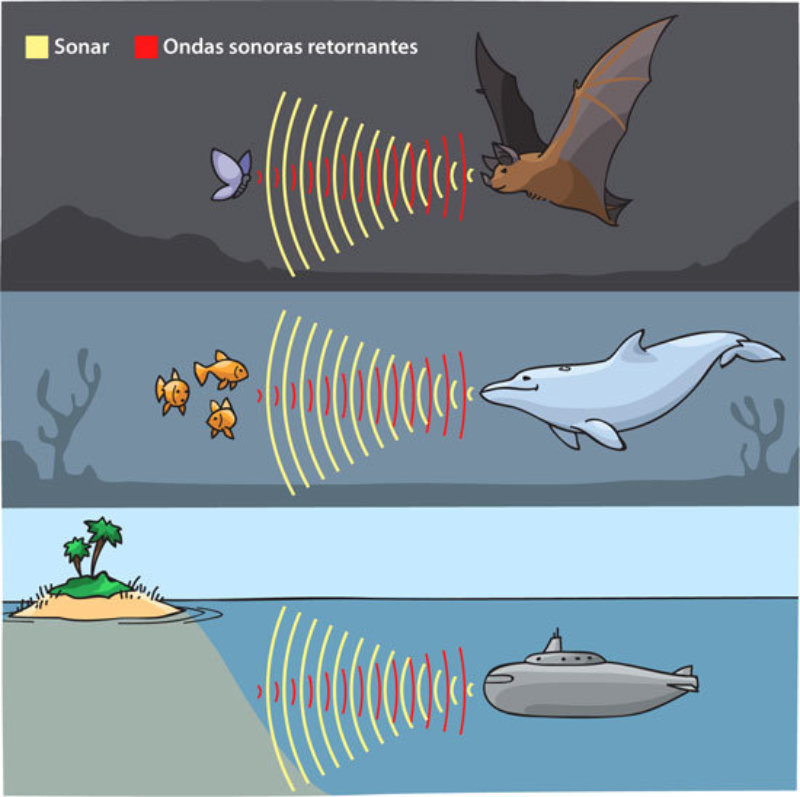
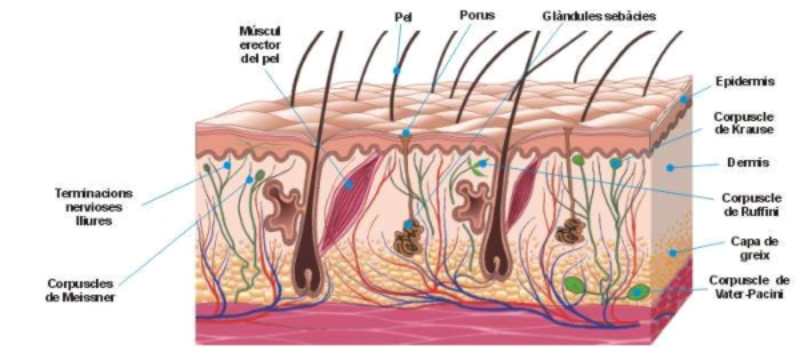
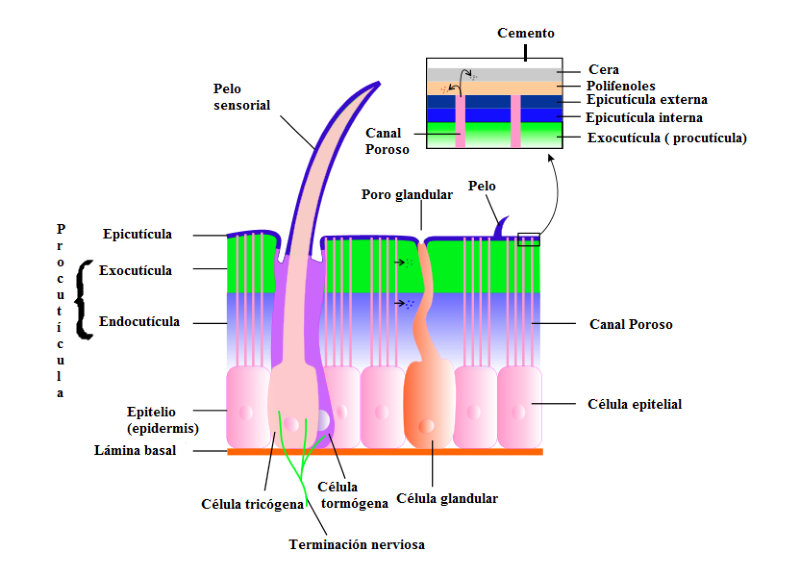
Perciben energía mecánica: presión (propiorreceptores y receptores táctiles), gravedad (estatorreceptores) o sonido (fonorreceptores). 3.1.1. PROPIORRECEPTORES Mecanorreceptores internos que informan de la contracción muscular, la tensión de los tendones, la posición y el movimiento de articulaciones. 3.1.2. RECEPTORES TÁCTILES En invertebrados, principalmente insectos, existen pelos táctiles repartidos por todo el cuerpo, llamados sensilias, que captan vibraciones y contacto. En vertebrados son mucho más complejos. Hay pelos táctiles (vibrisas), pero la mayoría de estos receptores están en la dermis y detectan frío, calor, dolor y presión, dando lugar al sentido del tacto. 3.1.3. ESTATORRECEPTORES Detectan cambios en la gravedad y, por tanto, determinan la posición del cuerpo y el equilibrio. En invertebrados el estatorreceptor es un saco llamado estatocisto. Este saco está lleno de líquido y revestido interiormente de células ciliadas sensoriales. En el interior hay una concreción calcárea llamada estatolito. Al moverse el animal, se mueve el estatolito, que presiona sobre las células ciliadas que mandan información sobre el cambio de posición. Suelen situarse en la cabeza (tentáculos, antenas,…). En vertebrados los estatorreceptores del equilibrio se localizan en el oído interno, dentro del sáculo y el utrículo. En ellos hay células ciliadas con estatolitos llamados otolitos. A su lado se hallan los canales semicirculares, órganos que informan del movimiento de la cabeza. Cada canal tiene en uno de sus extremos un ensanchamiento llamado ampolla con células ciliadas. 3.1.4. FONORRECEPTORES Son receptores del sonido, vibraciones y variaciones de presión en el aire o el agua. Los más importantes son el oído y la línea lateral. Oído: sólo presente en animales capaces de emitir sonido: artrópodos (crustáceos, arañas, insectos) y en los vertebrados. La localización en invertebrados es muy variable (antenas de mosquitos, patas de grillos, tórax de cigarras, abdomen de saltamontes, …). En los vertebrados se sitúa a los lados de la cabeza y es muy complejo. Suele tener un oído externo (pabellón auricular u oreja y conducto auditivo); oído medio (tímpano, cadena de huesecillos (martillo, yunque y estribo) y trompa de Eustaquio); y oído interno (caracol o cóclea, canales semicirculares, sáculo y utrículo). Línea lateral: aunque los peces tienen oído interno, sólo contiene los receptores del equilibrio (canales semicirculares, utrículo y sáculo) y carece de caracol, por lo que no funciona como fonorreceptor. Las vibraciones que forman el sonido las recibe mediante un órgano especial llamado línea lateral. Son tuberías de los costados del animal, llenas de líquido y con células sensoriales ciliadas que detectan las vibraciones del agua y la procedencia de las mismas. Ecolocación: algunos animales utilizan el sonido como un radar. Emiten sonidos cuyo eco les permite detectar presas u obstáculos incluso en la oscuridad. Es propio de murciélagos y delfines, por ejemplo. También lo tienen algunas aves que viven en cuevas (vencejos) y algunos mamíferos como el cachalote.3.2. TERMORRECEPTORES
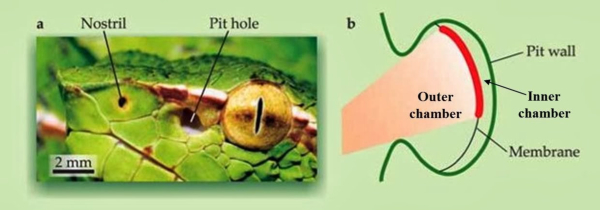
Detectan cambios de temperatura interna o externa. La respuesta puede ser un cambio fisiológico (homeostasis) o de comportamiento (letargo, migración, …). Algunos parásitos como las garrapatas detectan a sus huéspedes por el calor. También algunas serpientes presentan detectores de temperatura (foseta loreal) para detectar a sus presas de sangre caliente.3.3. MAGNETORRECEPTORES
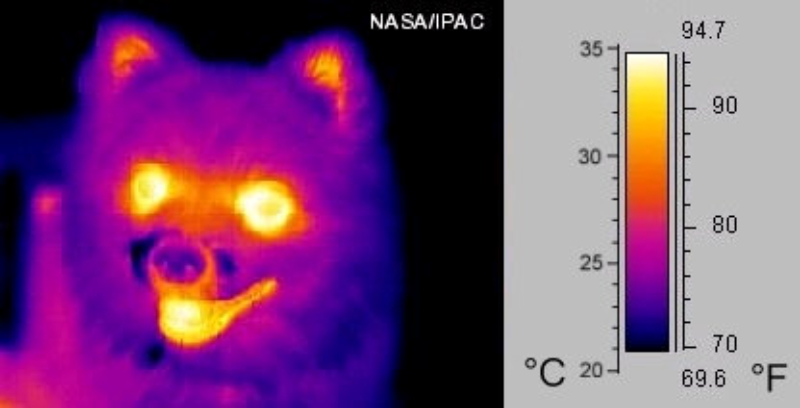
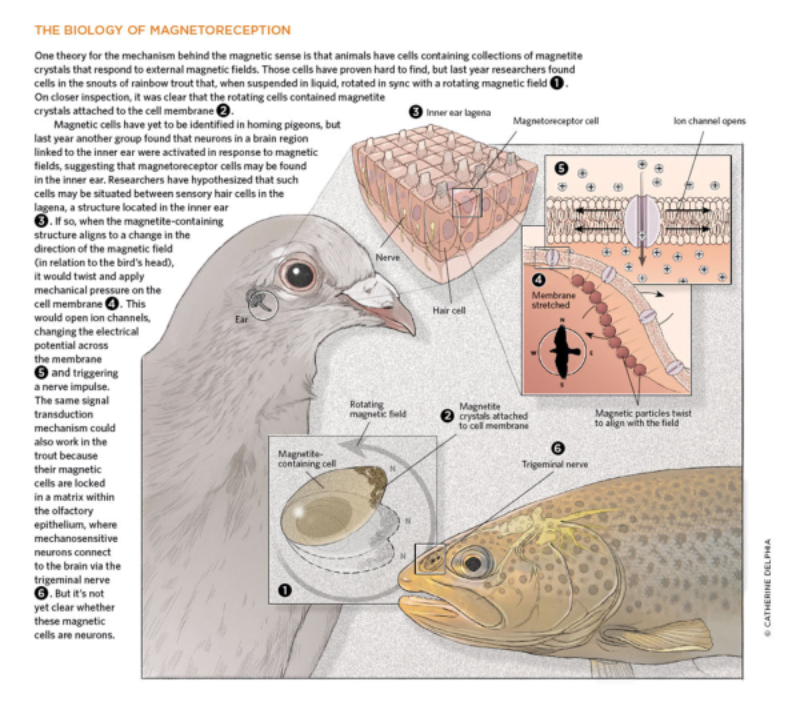
Detectan el campo magnético terrestre y permiten la orientación durante las migraciones. Abejas, tortugas marinas, algunas aves y delfines presentan cristales de magnetita que usan como brújulas.3.4. ELECTRORRECEPTORES
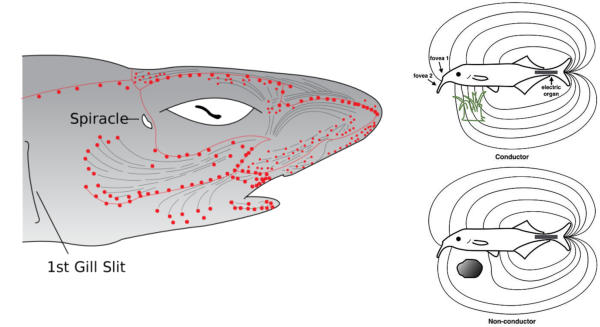
Receptores que perciben los campos eléctricos de músculos y nervios de otros animales. En los tiburones estos receptores se denominan ampollas de Lorenzini y les permite la comunicación y la detección de objetos en aguas con poca visibilidad.3.5. QUIMIORRECEPTORES
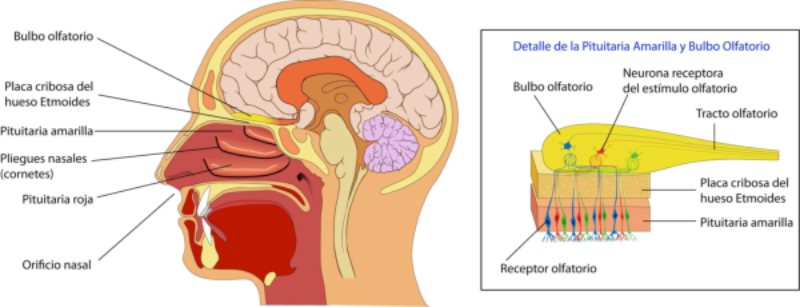
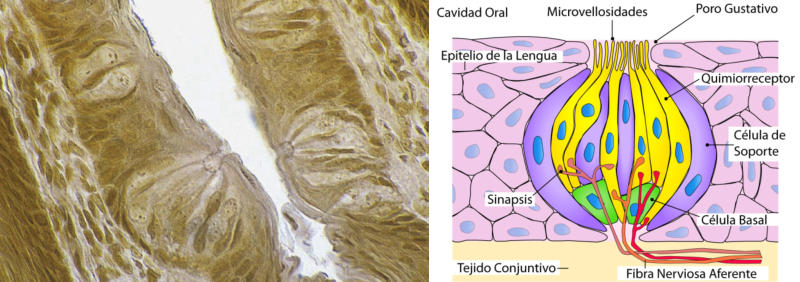
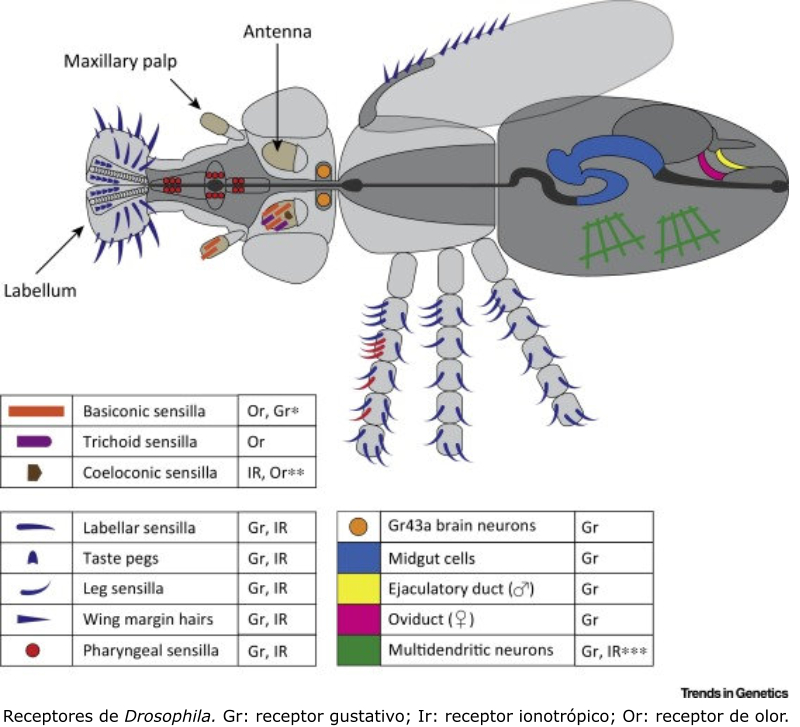
Detectan sustancias químicas disueltas o gaseosas. Son los receptores más primitivos, pues se relacionan con la nutrición y la reproducción. Los más importantes son los órganos del gusto y el olfato. Olfato: detecta sustancias químicas a distancia, tanto en el aire como en el agua. Es el principal sentido del comportamiento social. Los olores determinan un complejo lenguaje químico en muchos animales. Sirven para marcar territorios, establecer jerarquías, trazar senderos, atraer a la pareja sexual (feromonas), etc. Gusto: detecta sustancias químicas por contacto. Muchos invertebrados tienen este sentido en patas, antenas y otras partes del cuerpo. Los vertebrados lo tienen en la boca. En mamíferos se halla sobre todo en la lengua, donde los receptores forman papilas gustativas de distinta forma y especializadas en diferentes sabores.3.6. FOTORRECEPTORES
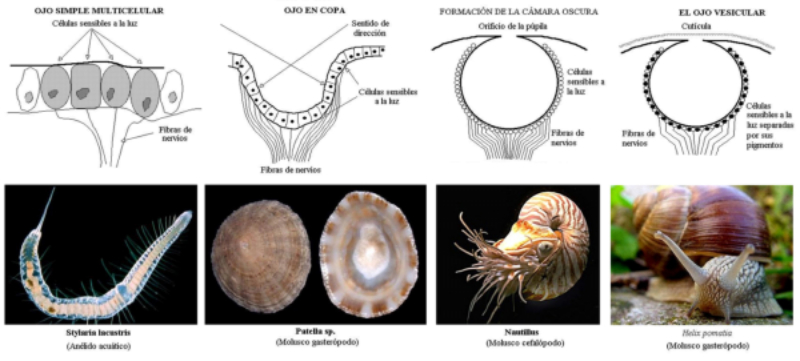
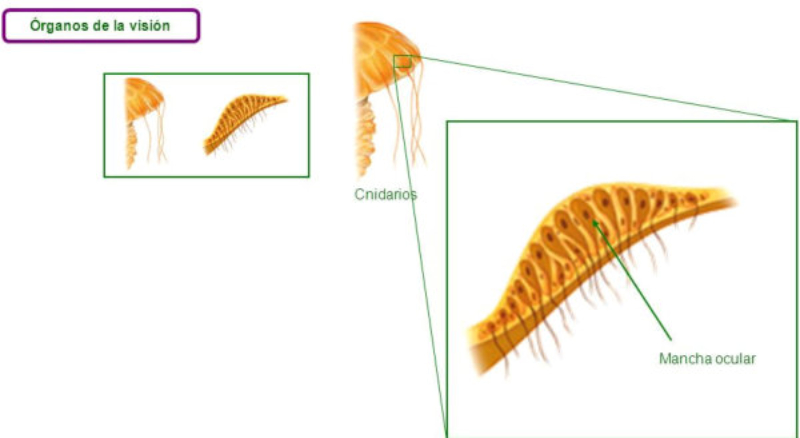
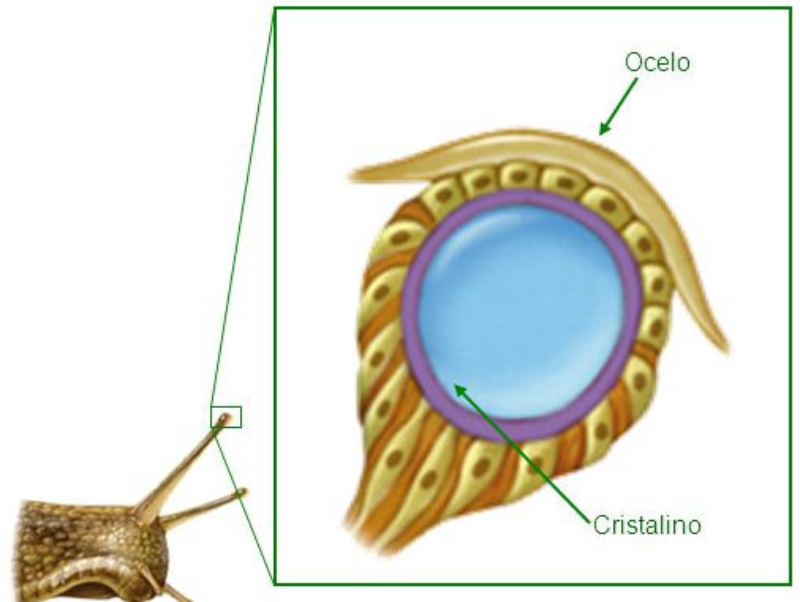
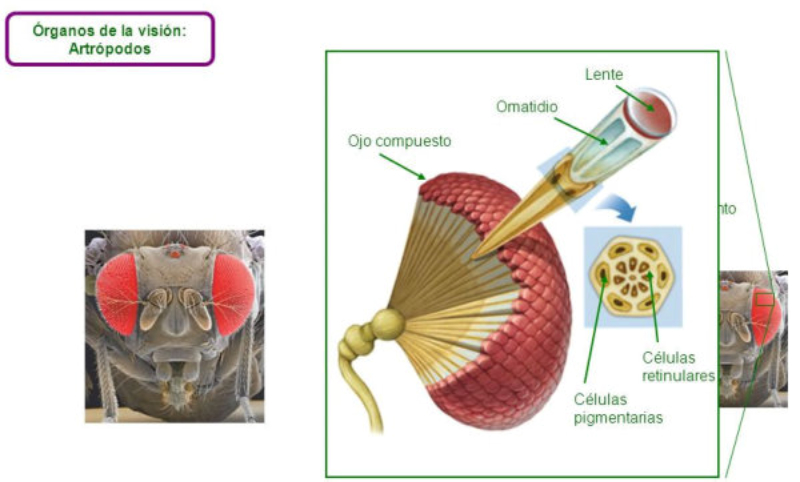
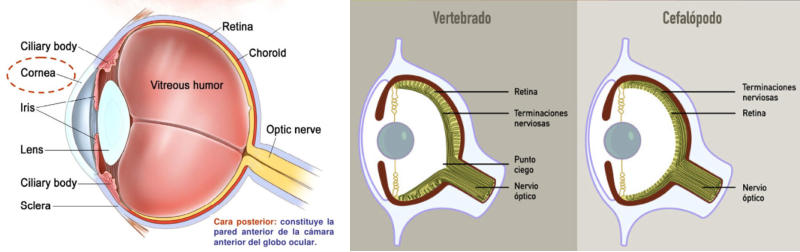
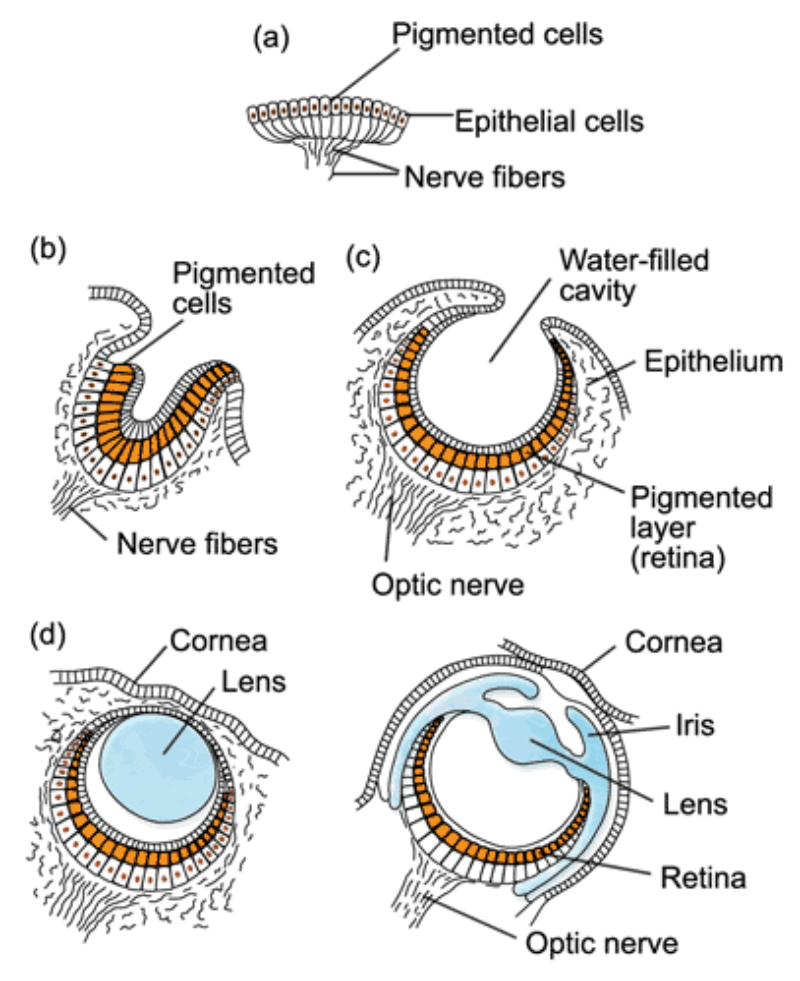
Los fotorreceptores son células sensibles a la luz gracias a pigmentos llamados rodopsinas. Estas células suelen agruparse formando la retina, la parte fotosensible del ojo u órgano de la visión. Básicamente, existen dos tipos de ojos: simples y compuestos. 3.6.1. OJOS SIMPLES Ocelo o mancha ocular: simples células sensibles en la superficie corporal. Típicos de cnidarios. Ojo en copa: la retina forma una depresión, lo que permite detectar la dirección de la luz. Carece de cristalino, por lo que no puede captar imágenes, sólo luces y sombras. Se halla en platelmintos (planarias). Ojo vesicular: tiene córnea y cristalino gelatinoso, por lo que puede enfocar las imágenes. Aparece en anélidos y gasterópodos. Ojo en cámara: el ojo es un globo con tres capas: esclerótica (con la parte anterior, la córnea transparente para el paso de luz); la coroides (vascularizada) y la retina. En la retina hay dos tipos de células fotosensibles: los bastones (visión nocturna en blanco y negro) y los conos (visión diurna en color: los hay para el rojo, el azul y el verde). Además, aparece un cristalino transparente (que actúa de lente para enfocar) y el iris (músculos que actúan como un diafragma, controlando la entrada de luz a través de un orificio central, la pupila). Aparece en vertebrados y en cefalópodos. 3.6.2. OJOS COMPUESTOS Aparecen en artrópodos. Están formados por multitud de unidades llamadas omatidios. cada omatidio capta una parte del campo visual del animal. Contiene una lente o faceta (formada por la córnea y un cono cristalino transparente). La lente concentra y dirige la luz a los fotorreceptores, llamados células retinianas. El cerebro integrará todas las imágenes de los omatidios para dar una imagen en mosaico, pixelada y con poca resolución, pero muy eficaz para detectar movimientos.






SISTEMAS NEUROENDOCRINO Y
LOCOMOTOR ANIMAL














Para ir a donde no se sabe hay que ir por donde no se sabe.” San Juan de la Cruz
“It must be a strange world not being a scientist, going through life not knowing--or
maybe not caring about where the air came from, where the stars at night came from
or how far they are from us. I WANT TO KNOW” Michio Kaku
“Nullius in verba” Robert Boyle, Christopher Wren y Robert Hooke













- Naturaleza básica de la vida
- Organización celular
- Organización pluricelular: animales
- Tejidos vegetales
- Diversidad: Moneras, Protoctistas, Hongos
- Diversidad: plantas y animales
- Digestivo animal
- Circulatorio animal
- Respiratorio y excretor animal
- Reproductor animal
- Neuroendocrino
- Nutrición en plantas
- Otras formas de nutrición. Movimientos
- Reproducción en plantas
- Historia de la Tierra
- Prácticas de laboratorio
- Vídeos
- Exámenes y trabajos
- Cálculo Nota Final














- Naturaleza básica de la vida
- Organización celular
- Organización pluricelular: animales
- Tejidos vegetales
- Diversidad: Moneras, Protoctistas, Hongos
- Diversidad: plantas y animales no cordados
- Diversidad: cordados
- Digestivo animal
- Circulatorio animal
- Respiratorio y excretor animal
- Reproductor animal
- Neuroendocrino
- Nutrición en plantas: fotosíntesis
- Otras formas de nutrición. Movimientos.
- Reproducción en plantas
- Historia de la Tierra
- Prácticas de laboratorio
- Vídeos
- Exámenes y trabajos
- Cálculo Nota Final


")
")


Batman Ends







Peces bioluminiscentes


")
")
![[x]](BG1BTO_htm_files/close.png "Close")