

BIOLOGÍA Y GEOLOGÍA




NUTRICIÓN VEGETAL
1. LA NUTRICIÓN DE LAS PLANTAS
Las plantas, a diferencia de animales y hongos, son organismos
autótrofos, que obtienen la materia orgánica mediante
fotosíntesis, es decir, utilizando energía lumínica. La obtención
de energía, en cambio, es similar a la de la mayoría de seres
vivos, empleando la respiración aerobia para degradar la
materia orgánica obtenida en la fotosíntesis.
Aunque todas las plantas tienen un metabolismo similar, existen
dos tipos de organización en las plantas con ciertas
peculiaridades nutritivas:
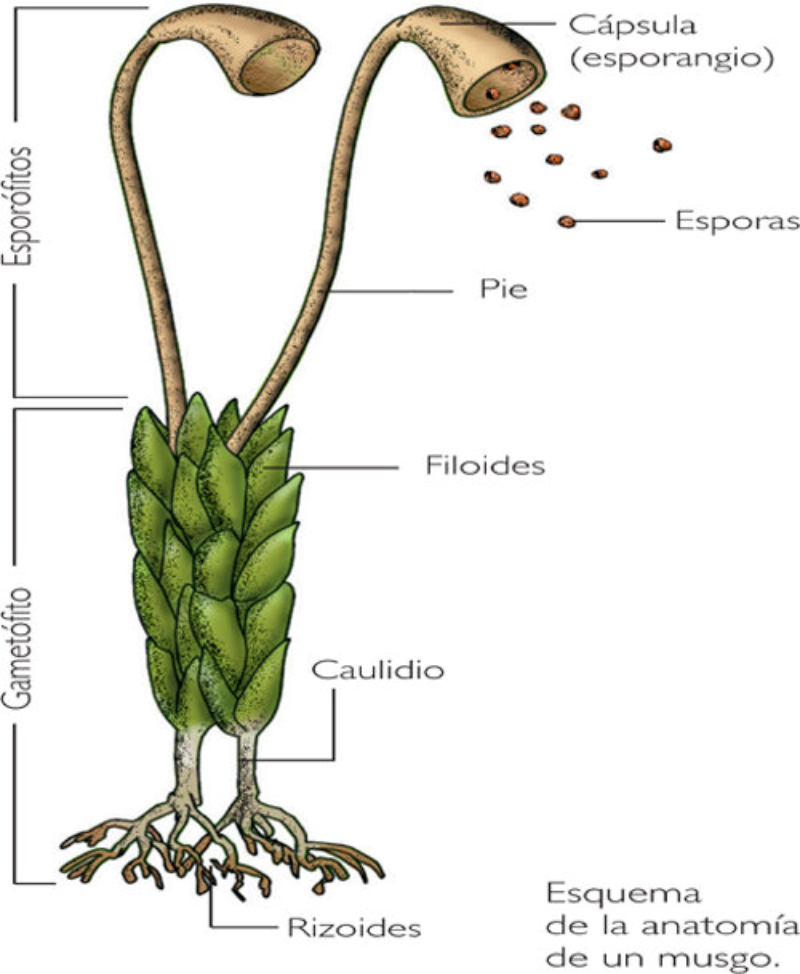
Plantas talofitas: tienen organización de tipo talo, es decir,
carecen de verdaderos tejidos y órganos. La absorción de
nutrientes y expulsión de desechos se realiza por toda la
superficie de la planta. El transporte es por difusión célula a
célula. La fotosíntesis se realiza en las partes verdes,
principalmente los filoides.
Plantas cormofitas: tienen organización de tipo cormo,
con verdaderos tejidos y órganos (raíz, tallo, hoja). Cada
órgano tiene un papel diferente en la nutrición: absorción de
nutrientes, intercambio de gases, transporte, fotosíntesis,
etc.
1.1. ABSORCIÓN DE NUTRIENTES POR LA RAÍCES
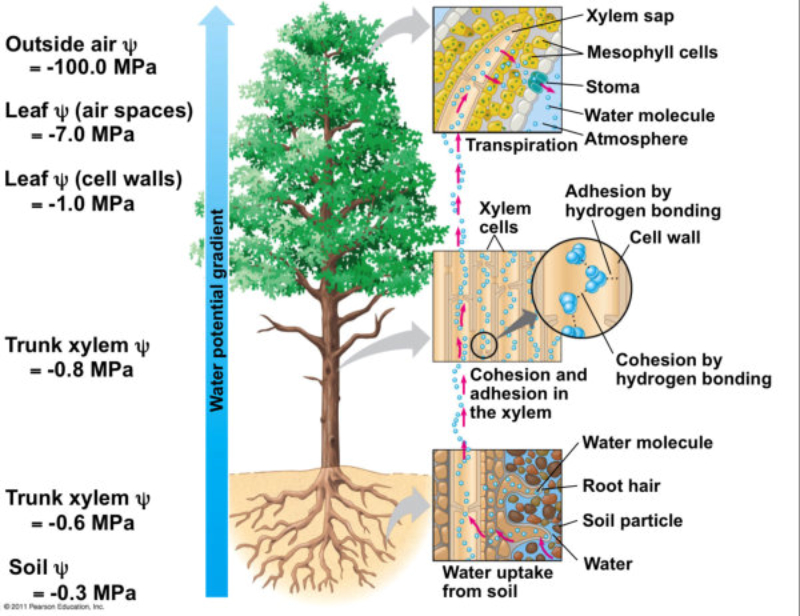
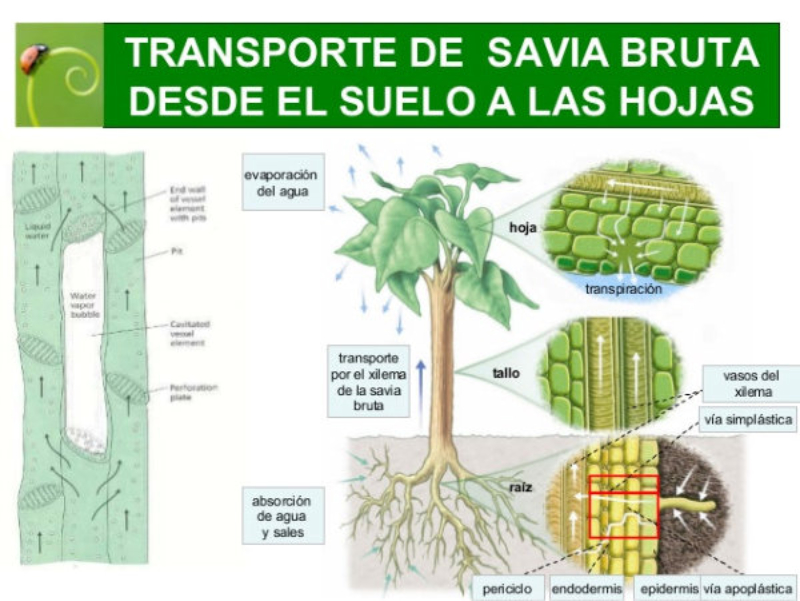
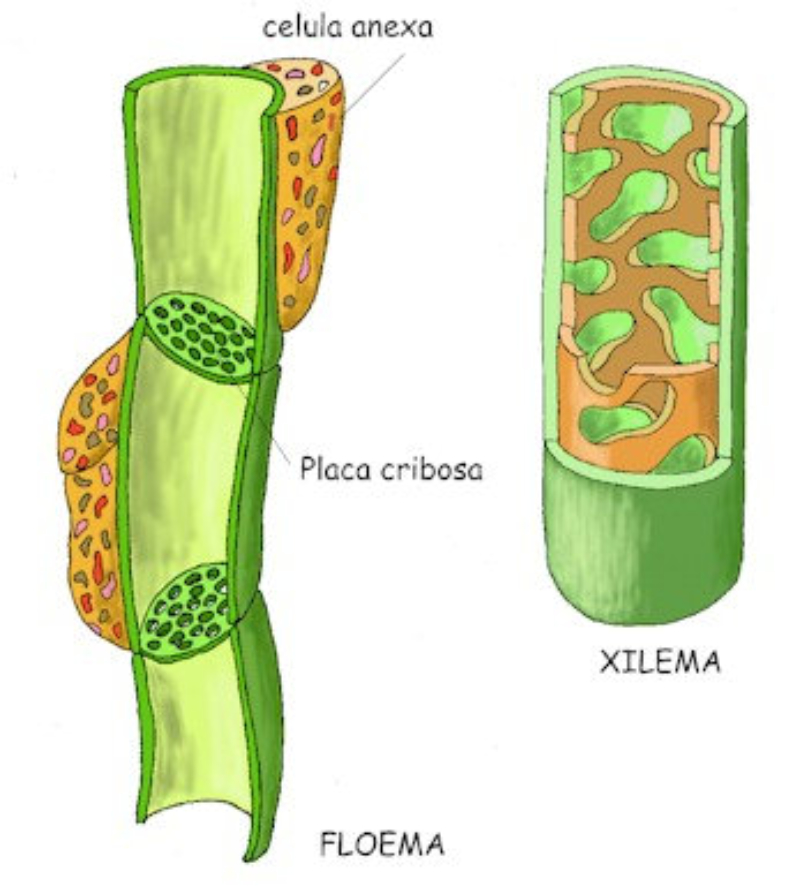
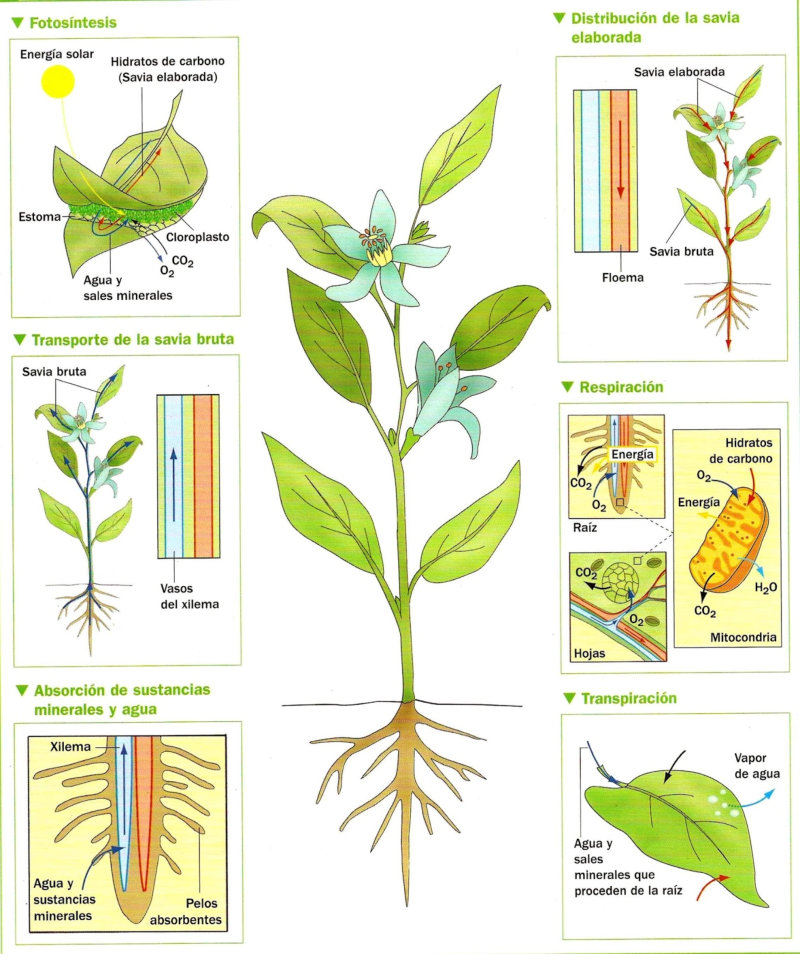
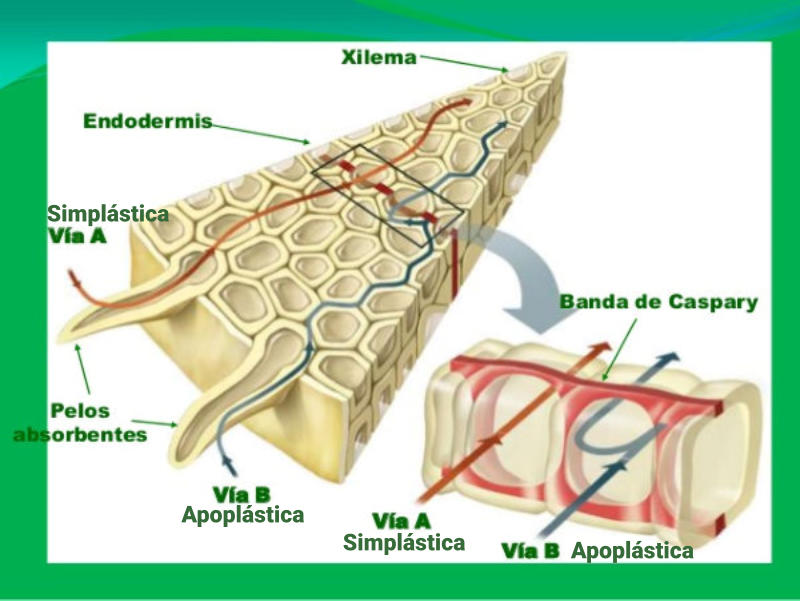
Las raíces, además de su papel de sostén de planta, permite la absorción de agua y sales minerales que, juntos, forman la savia bruta. La entrada de estos nutrientes se realiza por los pelos radicales y luego se dirigen hacia los vasos conductores del xilema para ser trasportados a las partes fotosintéticas. 1.1.1. ABSORCIÓN DE AGUA El agua entra en las células de la raíz por ósmosis, a favor de gradiente. Por lo que es un proceso pasivo que no requiere energía. Para lograrlo, las planta tiene una concentración de sales mayor en el citoplasma celular que en el agua del suelo. 1.1.2. ABSORCIÓN DE SALES MINERALES Las sales del suelo se hayan disueltas en forma de iones. Los iones atraviesan la membrana de las células epidérmicas de la raíz mediante transporte activo, contra gradiente y con gasto de energía. Una vez han entrado el agua y las sales en la rizodermis, debe llegar a los vasos conductores del xilema del centro de la raíz. Para ello, pueden seguir dos vías: Vía simplástica: atravesando el citoplasma celular y pasando de célula a célula a través de los plasmodesmos, orificios de la pared que comunican células contiguas. Vía apoplástica: el agua y las sales circulan en medio de las células, entre las paredes celulares, hasta que llegan a la endodermis. Allí, la banda de Caspary impide que sigan por esta vía, por lo que el agua y las sales entran en ellas células y siguen por la vía simplástica.1.2. TRANSPORTE DE LA SAVIA BRUTA
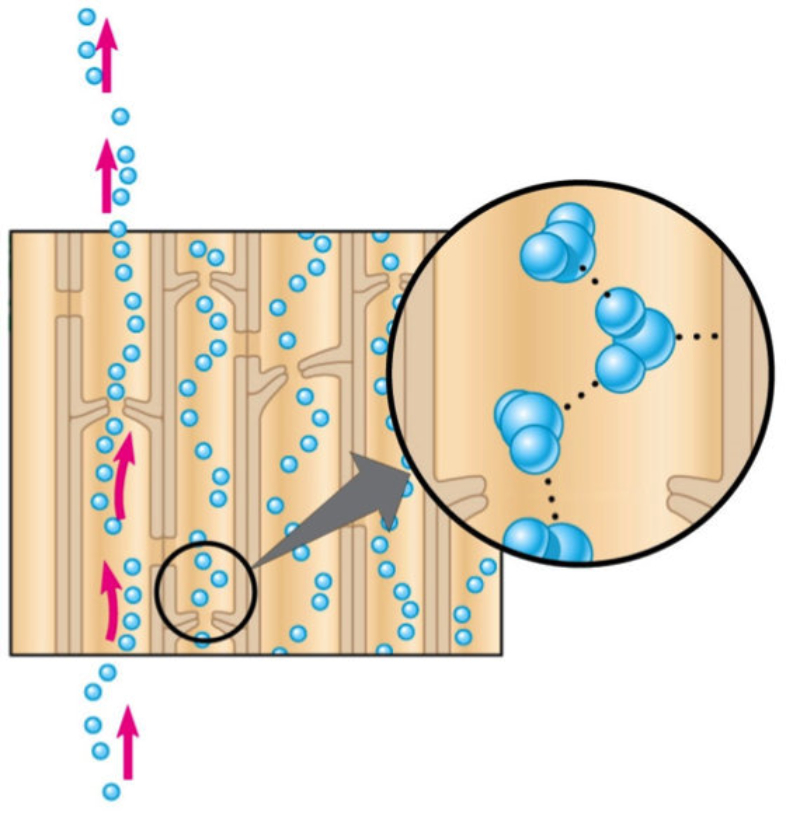
Una vez en el xilema, el agua y las sales deben ascender por los vasos leñosos hasta llegar a las zonas fotosintéticas, principalmente las hojas. Esto se consigue mediante la colaboración de tres mecanismos: la presión radicular y el mecanismo de cohesión-tensión, que incluye la transpiración y la capilaridad. 1.2.1. PRESIÓN RADICULAR La presión osmótica que impele al agua a entrar en las células de la raíz favorece el desplazamiento de la savia bruta de célula a célula y hacia arriba por el xilema. 1.2.2. MECANISMO DE COHESIÓN-TENSIÓN En plantas de gran tamaño se precisan otros procesos para el ascenso de la savia bruta que, juntos, forman el mecanismo de cohesión-tensión. Estos procesos son la transpiración y la capilaridad. TRANSPIRACIÓN La pérdida de agua a través de los estomas de las hojas provoca una presión negativa en los vasos leñosos, que se transmite de célula a célula y genera un efecto de succión sobre la savia bruta. CAPILARIDAD La capilaridad es la capacidad del agua de subir por tubos estrechos. Esta capacidad se debe a la adhesión de las moléculas de agua a las paredes de estos tubos y la cohesión entre moléculas de agua por los puentes de hidrógeno.1.3. INTERCAMBIO DE GASES
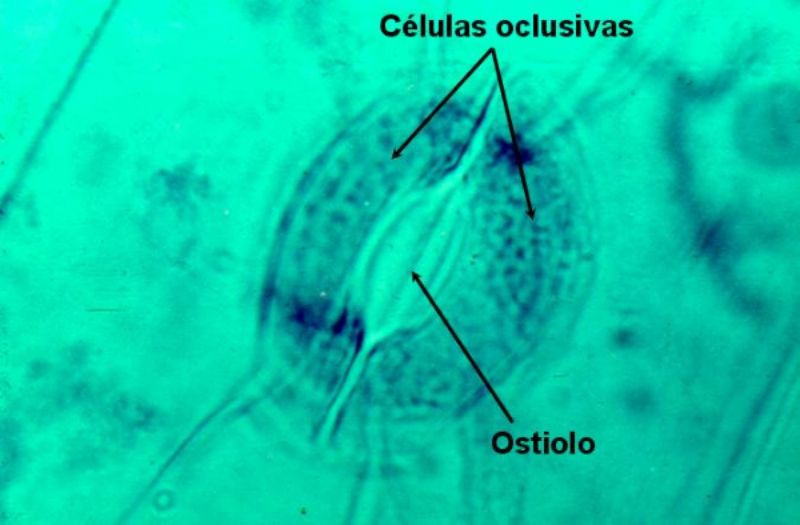
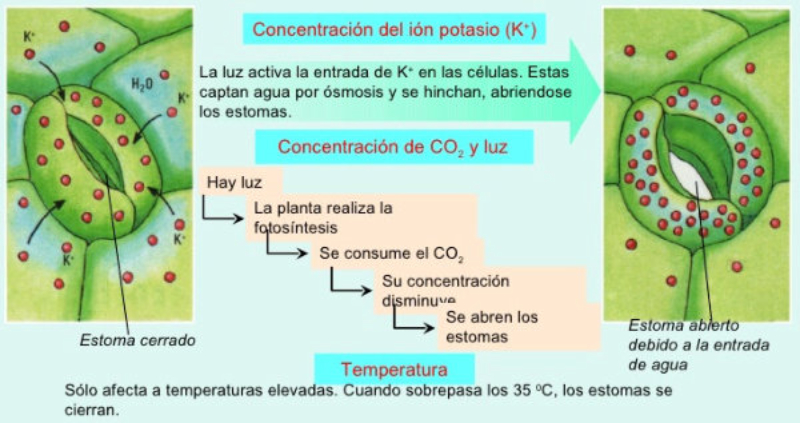
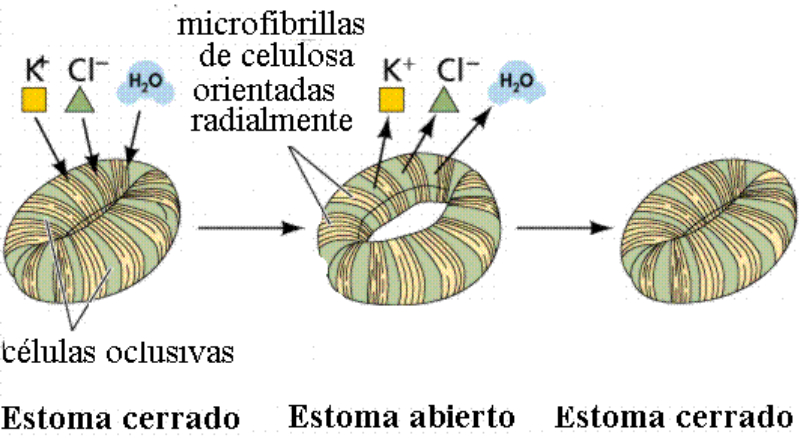
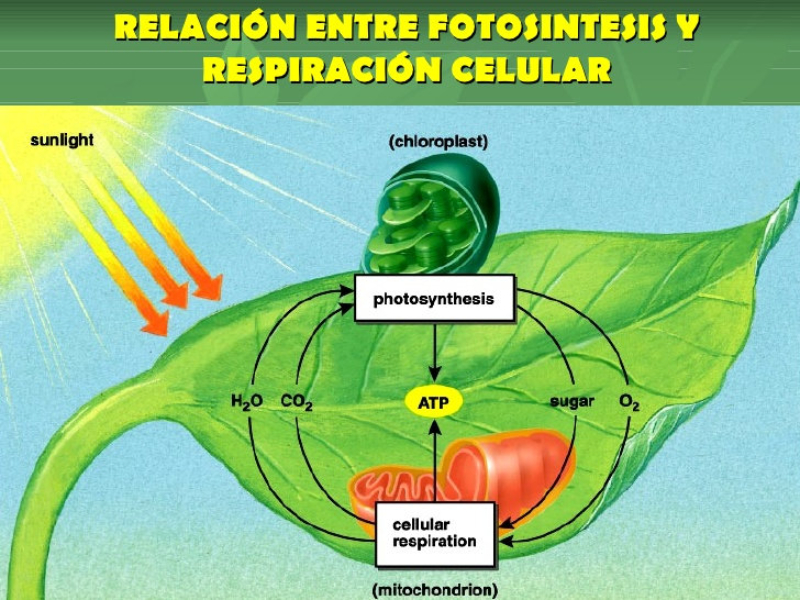
Las plantas precisan CO 2 para la fotosíntesis, en la que liberan O 2 . Por el contrario, en la respiración emplean O 2 y liberan CO 2 . Además, el agua sobrante es expulsada en forma de vapor de agua. El empleo de todos estos gases requiere mecanismos de intercambio de gases. Dicho intercambio se produce principalmente por los estomas, pero también por los pelos radicales y las lenticelas de la corteza. Al igual que en los animales, el intercambio de gases se produce de forma pasiva, por la difusión a favor de gradiente de concentración. 1.3.1. APERTURA Y CIERRE DE LOS ESTOMAS Los estomas están formados por dos células oclusivas de forma arriñonada que dejan un orificio en medio llamado ostiolo. Durante el día, las células oclusivas acumulan glucosa (debido a la fotosíntesis) e iones K + , volviéndose hiperosmóticas respecto a las células que las rodean. Esto hace que entre agua en las células estomáticas, se hinchen (turgencia) y se abra el estoma. Durante la noche se da el proceso contrario. Disminuye la concentración de glucosa y K + , la célula se vuelve hipotónica, sale agua y se cierran los estomas. La apertura y cierre de los estomas está controlada por diversos factores, entre ellos: La luz: favorece la apertura, pues activa la fotosíntesis y provoca la entrada de iones K + en las células oclusivas. La concentración de CO 2 : si es baja, los estomas se abren. Si es alta, se cierran. La temperatura: sólo afecta cuando se llega a un determinado límite. Por encima de cierta temperatura, muchas plantas cierran sus estomas para evitar pérdidas de agua.1.4. LA FOTOSÍNTESIS
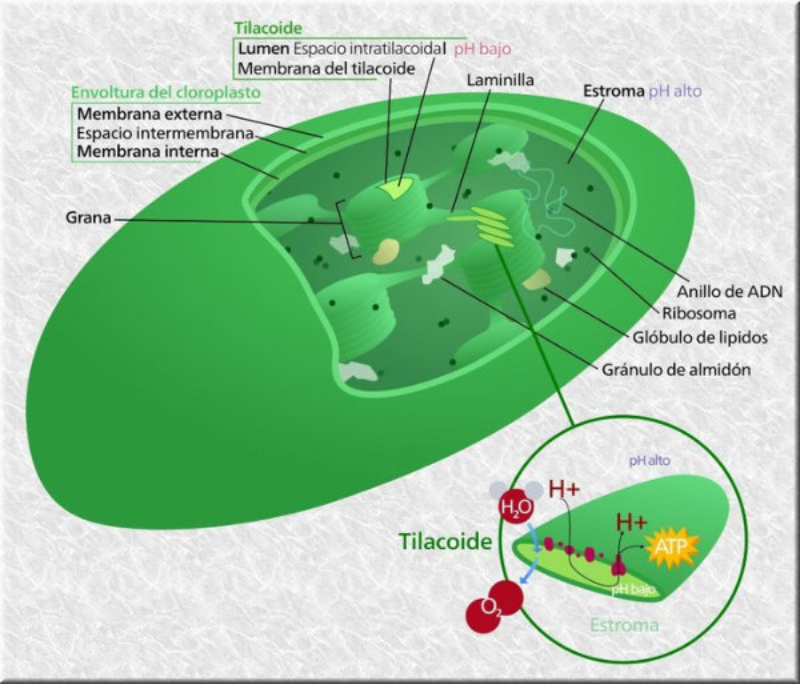
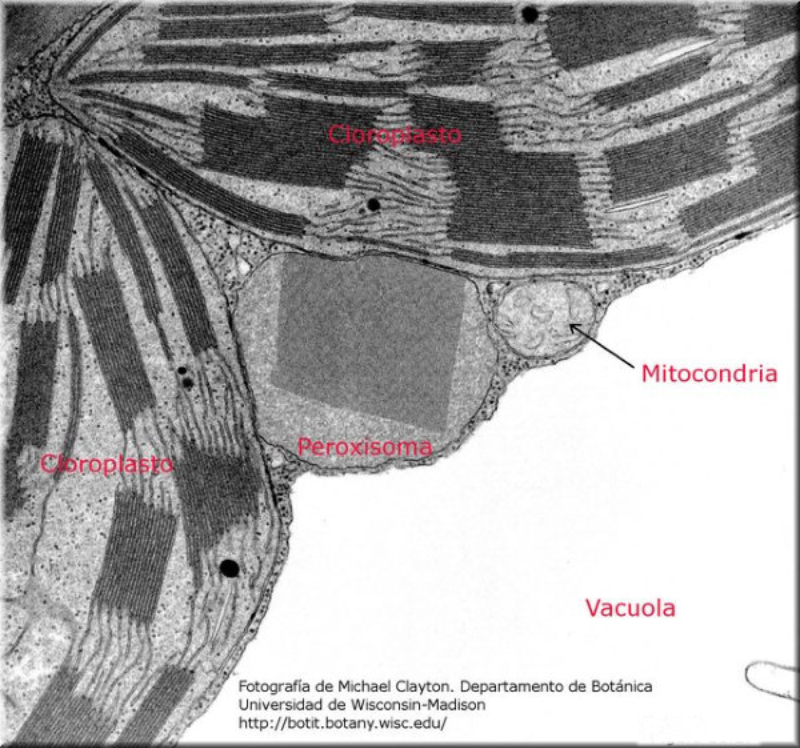
La fotosíntesis es un proceso anabólico autótrofo. La energía lumínica es empleada para obtener materia orgánica a partir de compuestos inorgánicos sencillos. La realizan las cianobacterias, algunas bacterias, las algas y las plantas. En las plantas la fotosíntesis se realiza en las partes verdes (tallos jóvenes y hojas), que presentan células con cloroplastos. 1.4.1. LOS CLOROPLASTOS Los cloroplastos son orgánulos membranosos exclusivos de las células eucariotas vegetales. Presentan tres tipos de membranas: Membrana externa: lisa y similar a la membrana plasmática, muy permeable. Membrana interna: también lisa, pero bastante impermeable, sobre todo a iones. Membrana tilacoidal: conjunto de sacos aplanados (tilacoides) en el interior del cloroplasto y donde se hallan los pigmentos fotosintéticos y, por tanto, se realiza la fotosíntesis. En algunas zonas los sacos del tilacoide aparecen apilados como monedas, formando los grana. Las tres membranas delimitan tres espacios en el cloroplasto: Espacio intermembranoso: entre la membrana externa e interna. Similar al citosol. Estroma: interior del cloroplasto, entre la membrana interna y la tilacoidal. Contiene una solución acuosa de sales, con proteínas, lípidos y glúcidos, enzimas, etc. También contiene ribosomas de tipo microbiano y cadenas de ADN circular de pequeño tamaño: el genoma del cloroplasto. Espacio tilacoidal: por dentro de la membrana tilacoidal. 1.4.2. FOTOSÍNTESIS La fotosíntesis es un proceso muy complejo y para su estudio se divide en dos fases: Fase lumínica: dependiente de la luz. Se realiza en los tilacoides y permite la captación de energía lumínica y su transformación en energía química (ATP y poder reductor). Fase oscura: no precisa de luz, aunque necesita los productos obtenidos en la fase anterior. Tiene lugar en el estroma y consiste en un conjunto de reacciones que transforma moléculas inorgánicas sencillas (CO 2 , H 2 O) en glucosa. FASE LUMÍNICA Transcurre en la membrana tilacoidal, donde la clorofila y otros pigmentos forman agrupaciones llamadas fotosistemas, que actúan como antenas captadoras de luz. Comprende tres procesos: Fotólisis del agua: la energía de los fotones de luz se emplea para romper moléculas de agua. Así se obtienen iones H + (que pasan al espacio tilacoidal y se emplearán en la fase oscura) y O 2 , que es liberado a la atmósfera. Reducción del NADP+: el NADP + (nicotín adenín dinucleótido fosfato) es un nucleótido capaz de transportar e - . Capta e- y H + y se convierte en NADPH, poder reductor. Síntesis de ATP: el ATP es un nucleótido rico en energía y la “moneda” energética de las células. Se forma en el proceso de fotofosforilación, gracias a la energía obtenida del transporte de e - . FASE OSCURA Sucede en el estroma. El ATP y el poder reductor (NADPH) obtenidos en la fase lumínica se emplean para reducir CO 2 a glucosa mediante reacciones que forman el ciclo de Calvin. Más tarde, también en el estroma, se emplearán diversas sales (P, N, S, …) para obtener lípidos, aminoácidos (para las proteínas) y nucleótidos (para los ácidos nucleicos). De esta forma, las células vegetales fabrican toda la materia orgánica. FACTORES QUE MODULAN LA FOTOSÍNTESIS El rendimiento de la fotosíntesis depende de numerosos factores ambientales, entre ellos: Intensidad lumínica: cada especie tiene un intervalo de luminosidad óptimo. Dentro de ese intervalo, el aumento de la luminosidad aumenta la fotosíntesis hasta un máximo. Concentración de CO 2 : a medida que aumenta, aumenta la actividad fotosintética, hasta llegar a un límite estable. Temperatura: en general, el aumento de la temperatura incrementa la fotosíntesis, pues aumenta la actividad enzimática. Por encima de cierto valor, la fotosíntesis disminuye hasta desaparecer por la desnaturalización de las enzimas. De todas formas, hay plantas adaptadas a climas cálidos y otras a climas fríos. Humedad: la falta de humedad provoca el cierre de los estomas para evitar la transpiración, lo que impide el intercambio de gases y la captación de CO 2 y lleva a la disminución de la actividad fotosintética.1.5. TRANSPORTE DE SAVIA ELABORADA
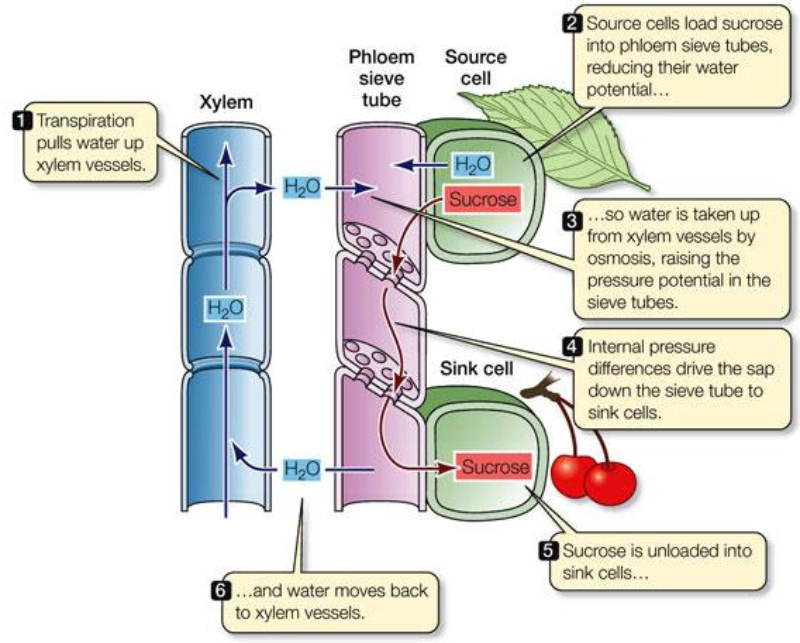
Los monosacáridos fabricados en la fotosíntesis se convierten en sacarosa y otras moléculas sencillas. La savia elaborada está formada por agua, sacarosa, aminoácidos, hormonas, proteínas, etc. El floema la llevará a donde se necesite. El transporte de la savia elaborada se denomina translocación y se produce entre las zonas fuente (lugares donde se fabrica:partes verdes fotosintéticas) y las zonas sumidero (donde se utiliza o almacena: raíces, tallos, frutos, flores,…). Las zonas sumidero, no obstante, pueden actuar como zonas fuente cuando la planta necesita sus reservas (tubérculos y raíces). Aunque el mecanismo no es del todo comprendido, la hipótesis más aceptada actualmente es la del flujo por presión. TRANSLOCACIÓN: HIPÓTESIS DEL FLUJO POR PRESIÓN Esta hipótesis propone que el transporte de la savia elaborada se produce gracias a una diferencia de presión entre las células de la fuente y las del sumidero. Dicha diferencia de presión se debe a procesos osmóticos. En las zonas fuente, la sacarosa pasa por transporte activo (con gasto de ATP) desde las células fotosintéticas a las del floema (tubos cribosos). Las células cribosas se vuelven así hipertónicas respecto a las vecinas del xilema. Esto provoca la entrada de agua desde el xilema al floema. Las células del floema se hinchan y sufren turgencia, aumentando su presión. La presión fuerza a la savia elaborada a desplazarse por las células contiguas del floema a través de las placas cribosas. El fenómeno se repite hasta llegar a la zona sumidero. En la zona sumidero la sacarosa sale del floema hacia las células de dicha zona mediante transporte activo. Esto disminuye la presión osmótica de las células floemáticas, lo que provoca que parte del agua que contienen pase al xilema contiguo que transporta savia bruta.1.6. USO DE LOS PRODUCTOS FOTOSINTÉTICOS
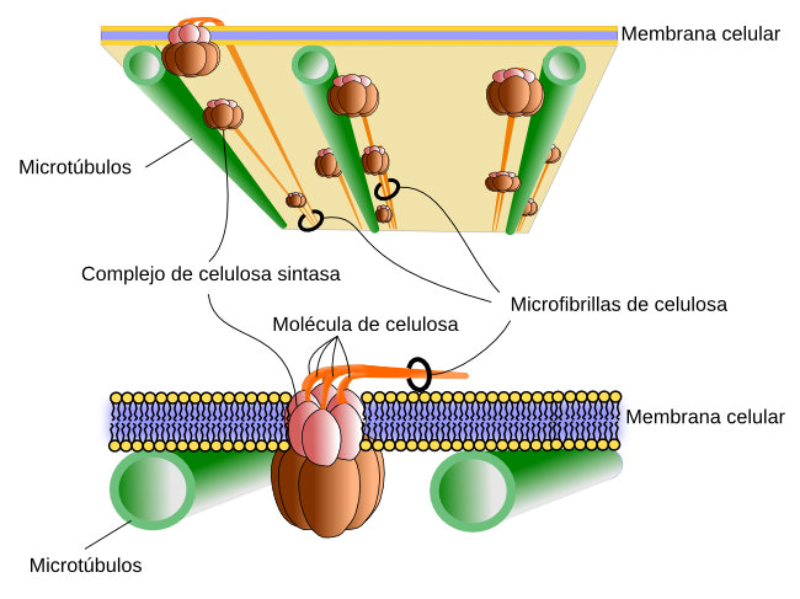
La mayor parte de la energía en forma de ATP producida en la fotosíntesis se emplea para sintetizar materia orgánica en la fase oscura. Dicha materia orgánica se empleará posteriormente en multitud de actividades de la planta: RESPIRACIÓN CELULAR Como la inmensa mayoría de seres vivos, las plantas emplean la respiración como principal proceso catabólico. La oxidación de glucosa y otras biomoléculas en el citosol y las mitocondrias genera energía en forma de ATP, CO 2 y H 2 O. SÍNTESIS DE CELULOSA La pared de las células vegetales está compuesta principalmente de celulosa. Este polisacárido se fabrica sobre la propia membrana plasmática gracias a enzimas que emplean glucosa y forman largas cadenas. RESERVA Las plantas producen más materia orgánica de la que precisan para sus propias funciones. El sobrante es almacenado como reserva de maneras variadas: Almidón: principal molécula de reserva vegetal, se almacena en raíces (zanahoria, remolacha), tallos (caña, tubérculos, bulbos) y semillas (cereales, legumbres). Lípidos: se emplea principalmente como reserva para los embriones en las semillas oleaginosas: aceitunas, frutos secos. Proteínas: se concentra sobre todo en el endospermo de las semillas, como fuente de aminoácidos para el embrión. También pueden hallarse en raíces y bulbos. OTRAS FUNCIONES Las plantas fabrican muchas otras sustancias con funciones específicas. Algunas se almacenan o se excretan. Esto incluye pigmentos (clorofilas, terpenos); esencias vegetales, derivados fenólicos (lignina, taninos) y alcaloides (drogas, medicamentos o venenos).1.7. EXCRECIÓN Y SECRECIÓN EN PLANTAS
Las sustancias obtenidas por las plantas que no le son de utilidad pueden almacenarse o bien ser expulsadas por excreción o secreción. Excreción: expulsión de sustancias sin utilidad para la planta. Las plantas carecen de órganos excretores especializados. Secreción: expulsión de sustancias que pueden realizar algún tipo de función como defensa. EXCRECIÓN EN PLANTAS Los gases, como el O 2 y el CO 2 se expulsan mediante el intercambio de gases. Otras sustancias han de ser excretadas: Cloruro sódico (NaCl): el exceso de sales en los suelos es un grave problema para las plantas. Sin embargo, las plantas halófitas se adaptan a suelos salinos expulsando el exceso de cloruro a través de glándulas especiales de las hojas. Oxalato cálcico (CaC 2 O 4 o Ca(COO) 2 ): esta sal no puede eliminarse y se acumula en vacuolas. Su ingesta en grandes cantidades (espinacas, apio, acelgas, aceitunas, frutas del bosque,…) puede provocar cálculos renales y otras alteraciones. SECRECIÓN EN PLANTAS Las plantas presentan numerosas glándulas secretoras que producen diversas sustancias con función básicamente defensiva: resinas, látex, néctar, esencias, alcaloides (quinina, cafeína, cocaína, estricnina), taninos, gomas.























Para ir a donde no se sabe hay que ir por donde no se sabe.” San Juan de la Cruz
“It must be a strange world not being a scientist, going through life not knowing--or
maybe not caring about where the air came from, where the stars at night came from
or how far they are from us. I WANT TO KNOW” Michio Kaku
“Nullius in verba” Robert Boyle, Christopher Wren y Robert Hooke
















- Naturaleza básica de la vida
- Organización celular
- Organización pluricelular: animales
- Tejidos vegetales
- Diversidad: Moneras, Protoctistas, Hongos
- Diversidad: plantas y animales
- Digestivo animal
- Circulatorio animal
- Respiratorio y excretor animal
- Reproductor animal
- Neuroendocrino
- Nutrición en plantas
- Otras formas de nutrición. Movimientos
- Reproducción en plantas
- Historia de la Tierra
- Prácticas de laboratorio
- Vídeos
- Exámenes y trabajos
- Cálculo Nota Final












- Naturaleza básica de la vida
- Organización celular
- Organización pluricelular: animales
- Tejidos vegetales
- Diversidad: Moneras, Protoctistas, Hongos
- Diversidad: plantas y animales no cordados
- Diversidad: cordados
- Digestivo animal
- Circulatorio animal
- Respiratorio y excretor animal
- Reproductor animal
- Neuroendocrino
- Nutrición en plantas: fotosíntesis
- Otras formas de nutrición. Movimientos.
- Reproducción en plantas
- Historia de la Tierra
- Prácticas de laboratorio
- Vídeos
- Exámenes y trabajos
- Cálculo Nota Final













Pelos radicales

Digitalis purpurea
Gutación (expulsión de agua del xilema) por
hidátodos (estomas fijos) en hojas de fresa.
Cristales de sal sobre una hoja.
Resina de conífera.





![[x]](BG1BTO_htm_files/close.png "Close")





