BIOLOGÍA Y GEOLOGÍA
LA CÉLULA: MORFOLOGÍA CELULAR
8. SISTEMAS INTERNOS DE MEMBRANAS
Además del núcleo, las células eucariotas presentan en su citoplasma orgánulos rodeados de membrana. En ellos se realizan actividades metabólicas especializadas y permite que células muy grandes funcionen de forma eficiente.8.1. RETÍCULO ENDOPLÁSMICO
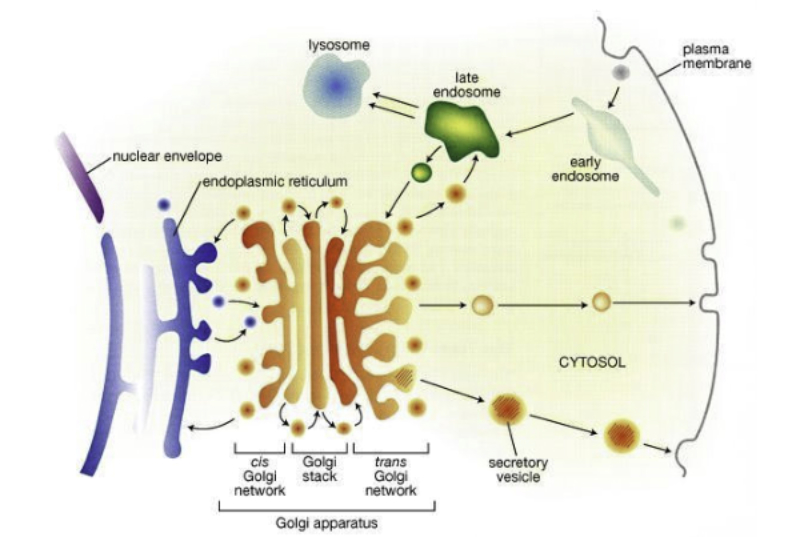
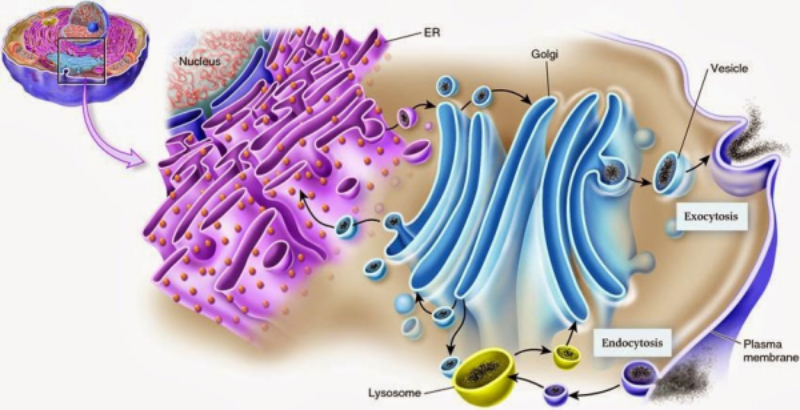
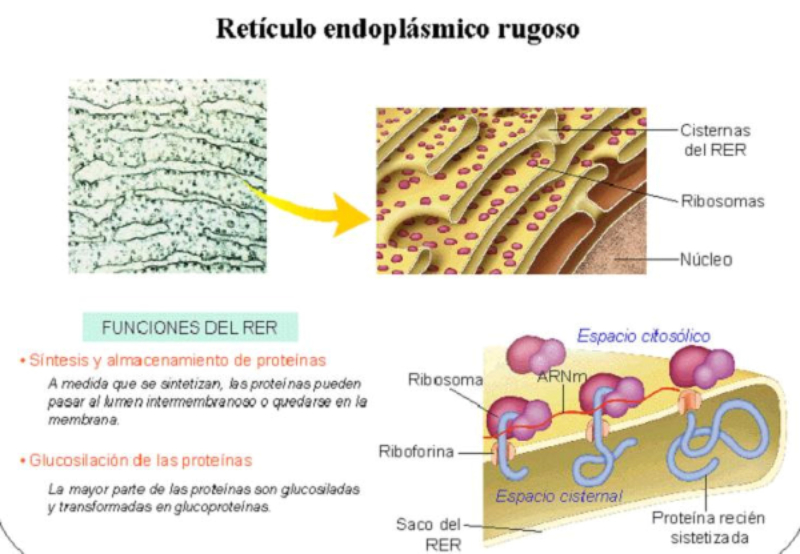
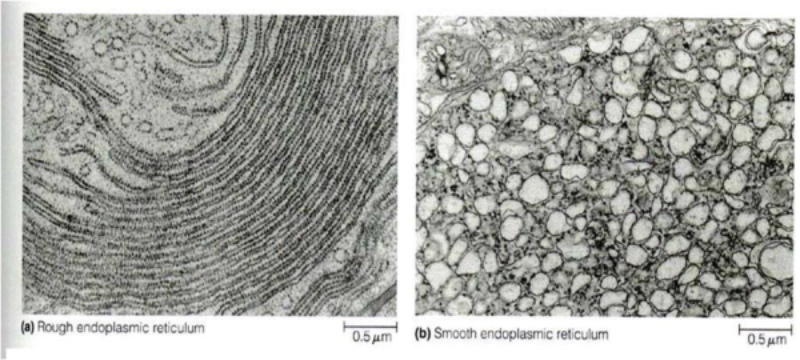
El retículo endoplásmico (RE) es un conjunto de sáculos aplanados y túbulos ramificados. Se extiende por todo el citoplasma y delimita un espacio cerrado y continuo que se comunica con el espacio perinuclear, pues el RE es continuo con la membrana nuclear externa. Es el mayor orgánulo en la mayoría de eucariotas. Existen dos tipos de RE: el retículo endoplásmico liso (REL) y el retículo endoplásmico rugoso (RER). REL: es un conjunto de túbulos lisos, sin ribosomas, con las siguientes funciones: Síntesis de lípidos y sus derivados: síntesis de fosfolípidos y colesterol En algunas células especiales se sintetizan hormonas esteroideas o ácidos biliares. Destoxificación de sustancias tóxicas liposolubles: las enzimas destoxificantes del REL son capaces de convertir sustancias tóxicas liposolubles (medicamentos, etanol, insecticidas, herbicidas, …) en hidrosolubles, excretables con la orina. Este proceso se realiza sobre todo en el hígado, pero también en intestino, riñones, piel y pulmones. Regulación del Ca 2+ intracelular: la membrana del REL presenta bombas de Ca 2+ y canales de Ca 2+ para la entrada y salida, respectivamente, de iones de Ca 2+ , lo que permite regular la secreción, la proliferación celular y las contracciones musculares. RER: también llamado ergastoplasma o retículo endoplásmico granular Formado por sáculos aplanados con numerosos ribosomas en su cara citosólica. Sus funciones son: Síntesis de proteínas: las que se van a secretar y las que formarán parte de la mayoría de los orgánulos (RE, Golgi, lisosomas y membrana plasmática). Inicio de la glucosilación: se le añaden los primeros azúcares. La glucosilación se terminará en el Golgi. Plegamiento de las proteínas fabricadas.8.2 APARATO DE GOLGI
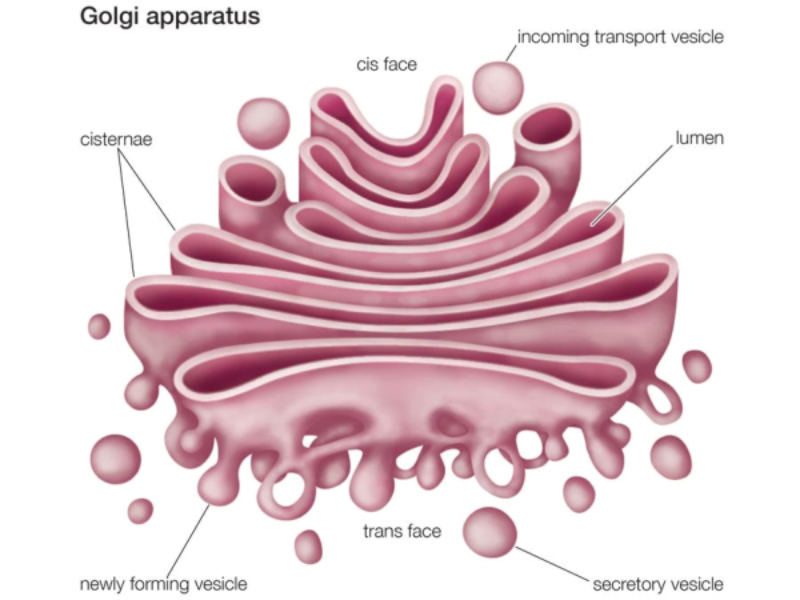
Conjunto de sáculos aplanados y apilados, con los extremos ensanchados, y numerosas vesículas acompañantes. Las pilas de sáculos, entre 5 y 8, interconectados entre sí, se denominan dictiosomas. Cada dictiosoma consta de tres regiones: Cara cis o de formación: cara cercana al núcleo. Convexa vista desde la membrana. Cara media: parte central. Cara trans o de maduración: cercana a la membrana plasmática y cóncava vista desde ésta. Las proteínas y lípidos sintetizados en el RE salen de éste en forma de vesículas de transporte que llegan a la cara cis. De aquí pasan a la cara media, donde sufren la mayor parte de las transformaciones, y terminan en la cara trans. Esta cara emite vesículas de transporte con destino a formar lisosomas, al exterior celular o a formar parte de la membrana plasmática. Funciones del aparato de Golgi Terminar la glucosilación de proteínas iniciada en el RER. Se da en las tres caras. Síntesis de glucolípidos y esfingomielina. Que formarán parte de la membrana plasmática. Síntesis de algunos polisacáridos de la pared celular en células vegetales. Aunque la celulosa se sintetiza sobre la propia membrana celular, las hemicelulosas y la pectina se fabrican en el Golgi y se transportan a la superficie. Distribución y exportación de proteínas. Salvo las proteínas del propio aparato de Golgi, todas las demás llegan a la cara trans. Allí se clasifican y se incluyen en diferentes vesículas según su destino final. Todas las células forman vesículas para dar lugar a lisosomas o formar parte de la membrana plasmática. Sin embargo, las células secretoras, forman vesículas especiales (con hormonas, neurotransmisores, enzimas digestivas,…) que quedan en el citoplasma y se secretan sólo en respuesta a estímulos.8.3. LISOSOMAS
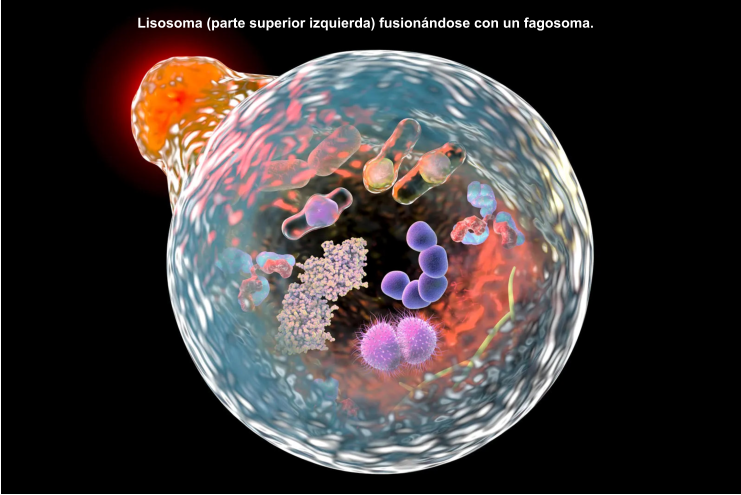
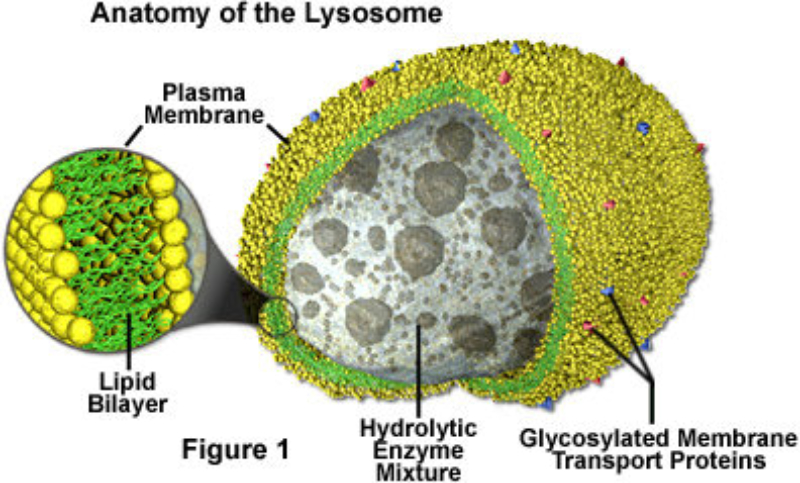
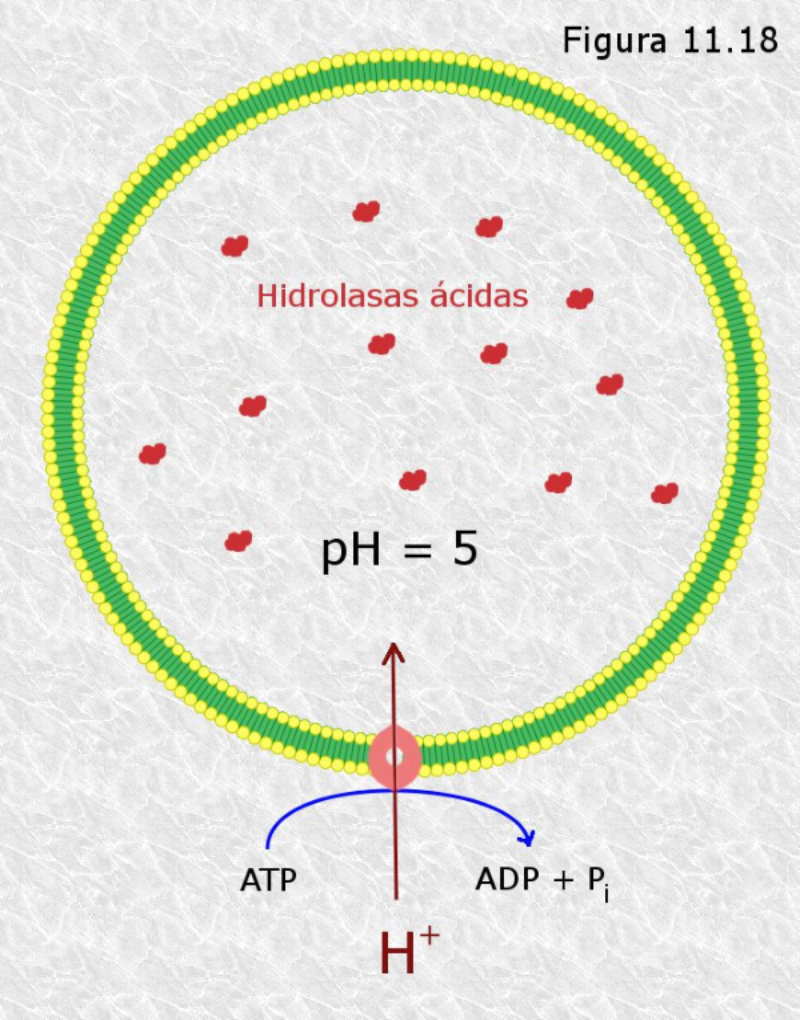
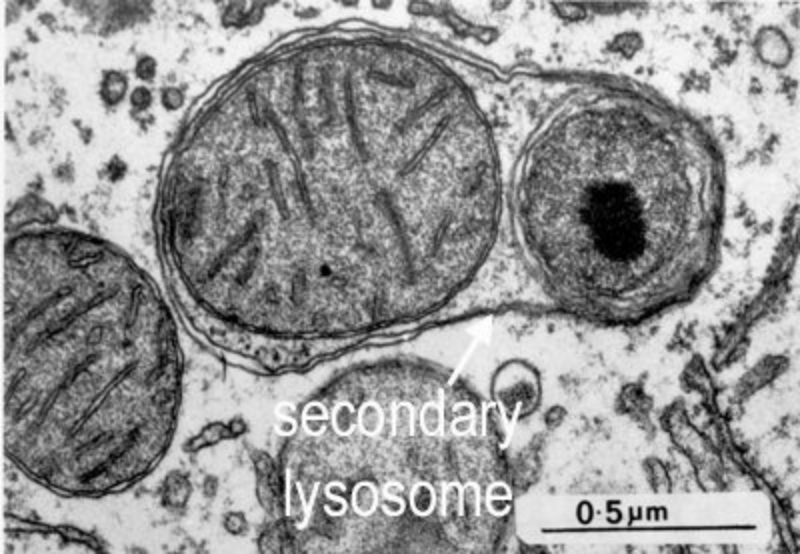
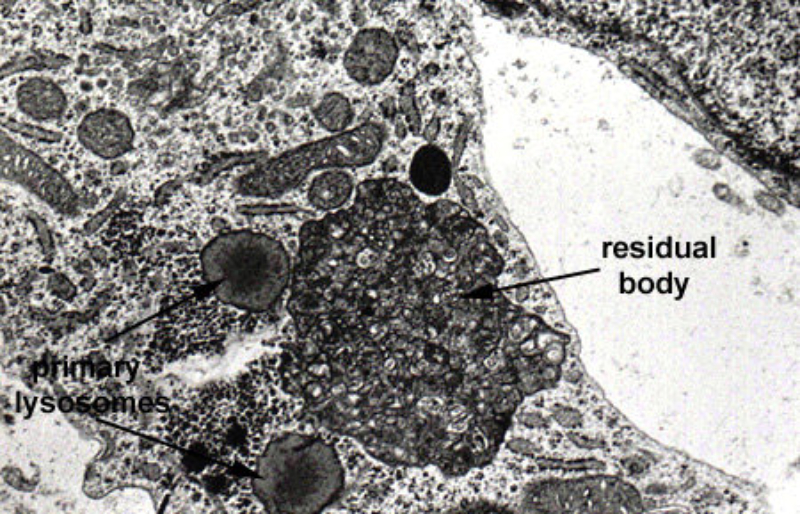
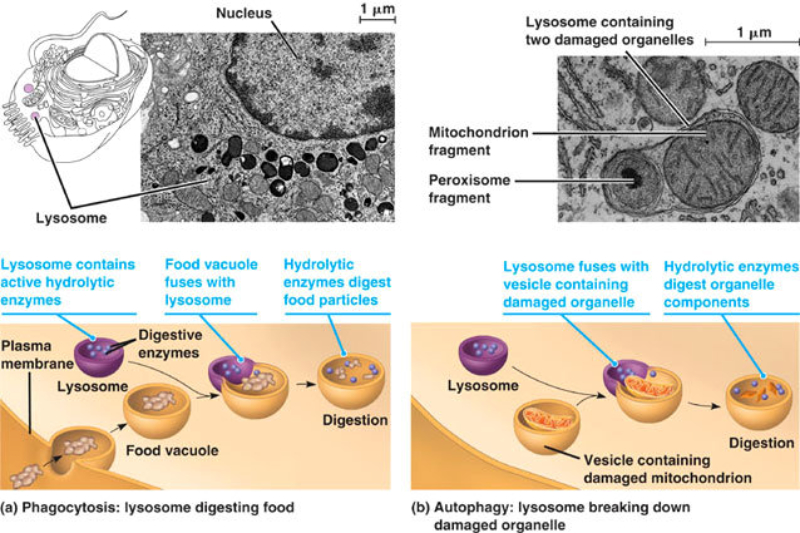
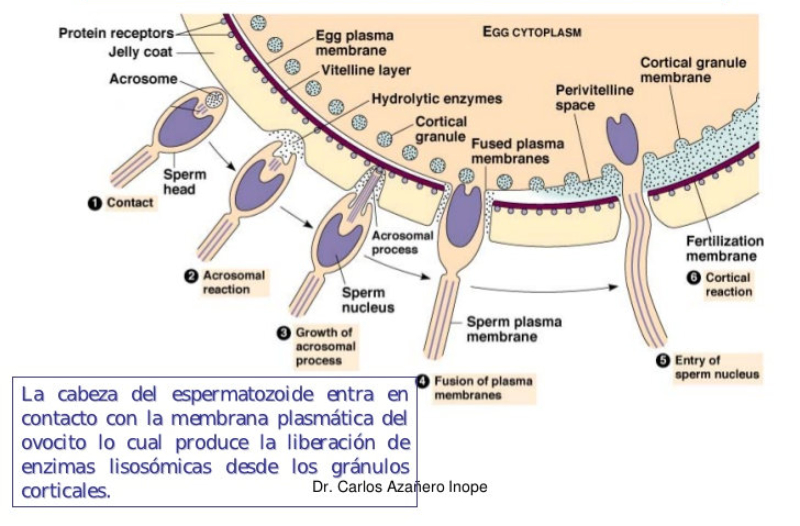
Los lisosomas son vesículas membranosas procedentes del aparato de Golgi que contienen enzimas hidrolíticas encargadas de la digestión intracelular de todo tipo de biomoléculas. Pueden digerir sustancias externas a la célula (heterofagia) o componentes celulares (autofagia). Las enzimas hidrolíticas de los lisosomas son todas hidrolasas ácidas, por lo que precisan un pH ácido para actuar. El interior del lisosoma debe estar a pH 5, por lo que dispone de bombas de protones que introducen H + en contra de gradiente. Además, la membrana del lisosoma contiene proteínas transportadoras para expulsar al citosol los productos de la digestión. En realidad, los lisosomas presentan gran variedad morfológica según el proceso digestivo en el que se encuentren: Lisosomas primarios: lisosomas que acaban de surgir del Golgi. Sólo contienen enzimas hidrolíticas y su contenido es homogéneo. Lisosomas secundarios: se forman al fusionarse los primarios con vesículas digestivas. Son de mayor tamaño y de contenido heterogéneo. Cuerpos residuales: lisosomas que han terminado el proceso digestivo. Son de tamaño variable y contienen sustancias no digeribles.8.4. PEROXISOMAS
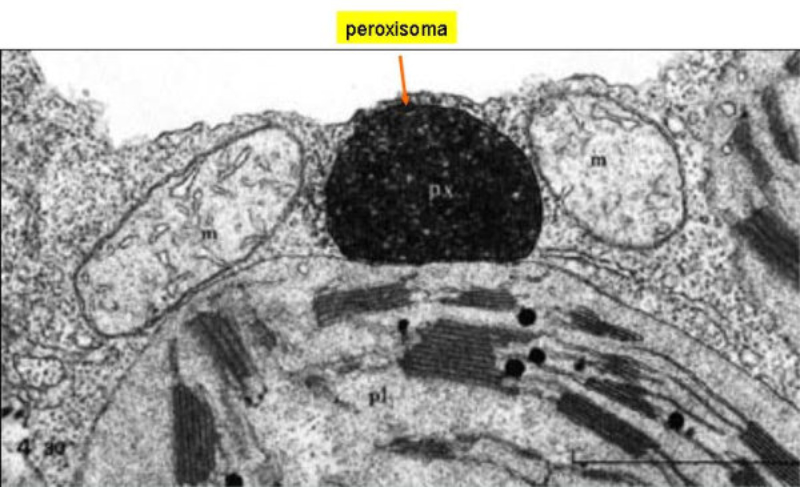
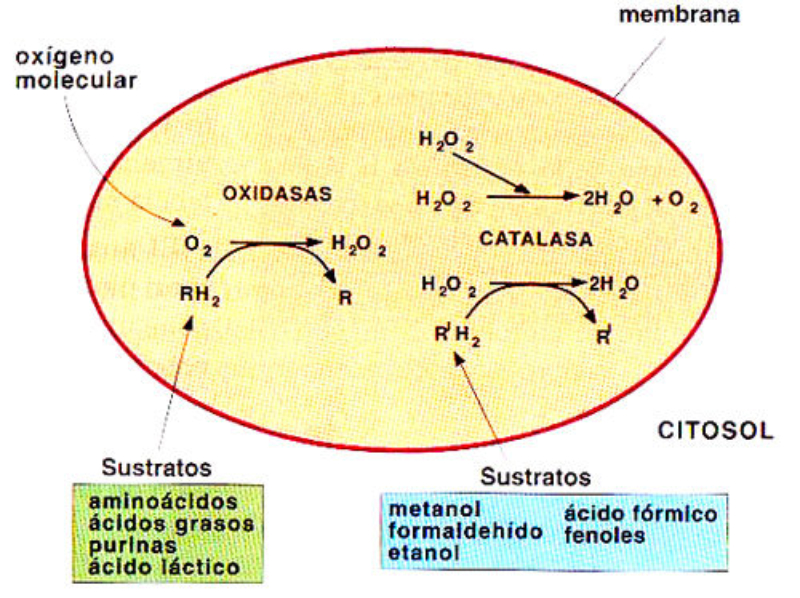
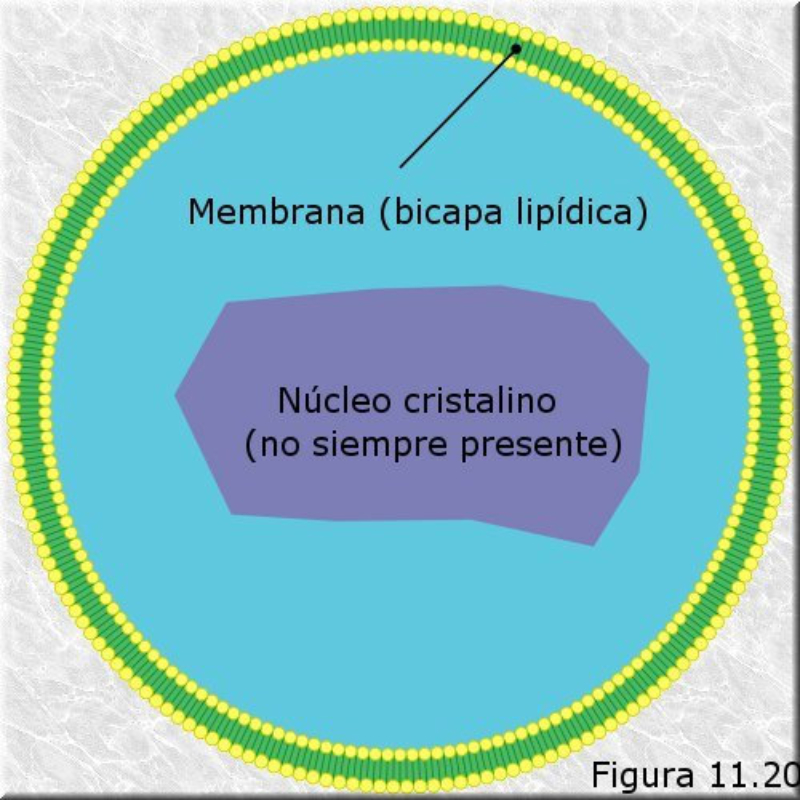
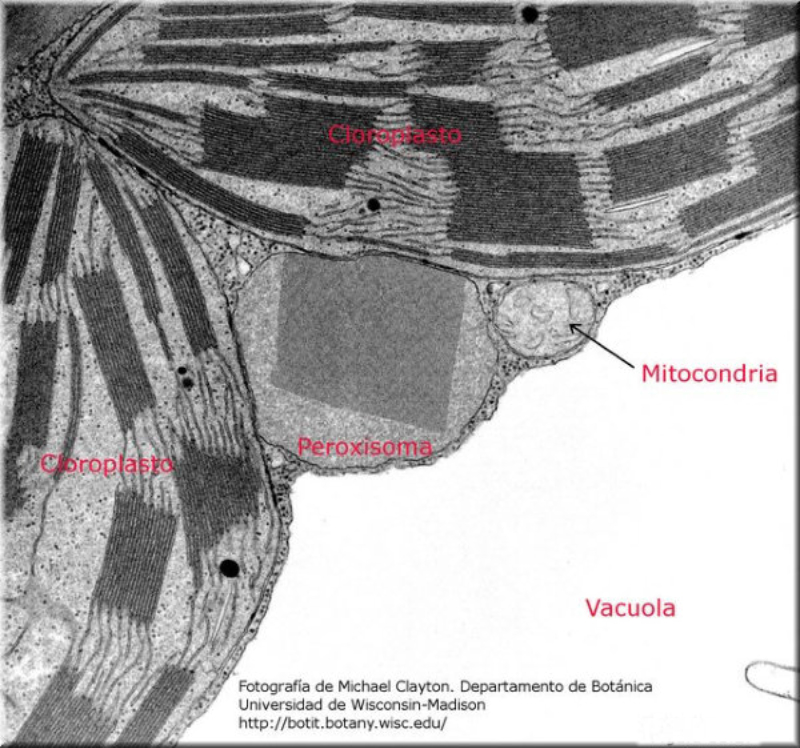
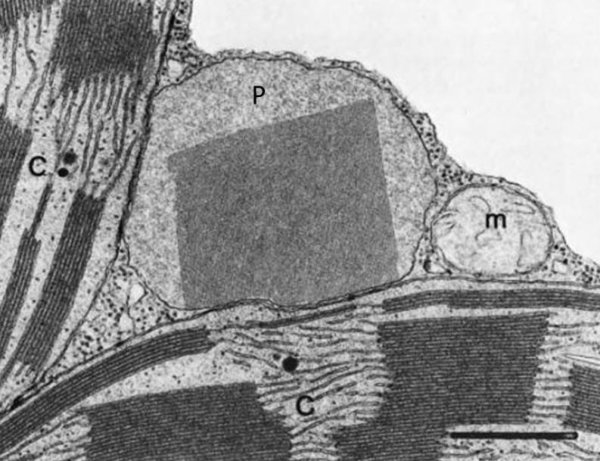
Vesículas membranosas de pequeño tamaño que contienen oxidasas, enzimas que utilizan el oxígeno molecular para oxidar compuestos orgánicos, obteniendo peróxido de hidrógeno (H 2 O 2 ). RH 2 + O 2 R + H 2 O 2 Dado que el peróxido de hidrógeno es muy tóxico para las células, los peroxisomas utilizan la enzima catalasa para degradarlo a agua, al tiempo que se oxidan sustancias como metanol, etanol, fenol,… H 2 O 2 + R’H 2 R’ + 2H 2 O Ambos procesos forman parte de la destoxificación que se dan en hígado y riñones principalmente. Además, los peroxisomas intervienen en el metabolismo lipídico de varias formas: Comienza la ß-oxidación de los ácidos grasos, que terminará en las mitocondrias. Síntesis de ácidos biliares a partir del colesterol. Síntesis de fosfolípidos y triglicéridos. Síntesis de isoprenoides. Fotorrespiración en plantas.8.5. MITOCONDRIAS
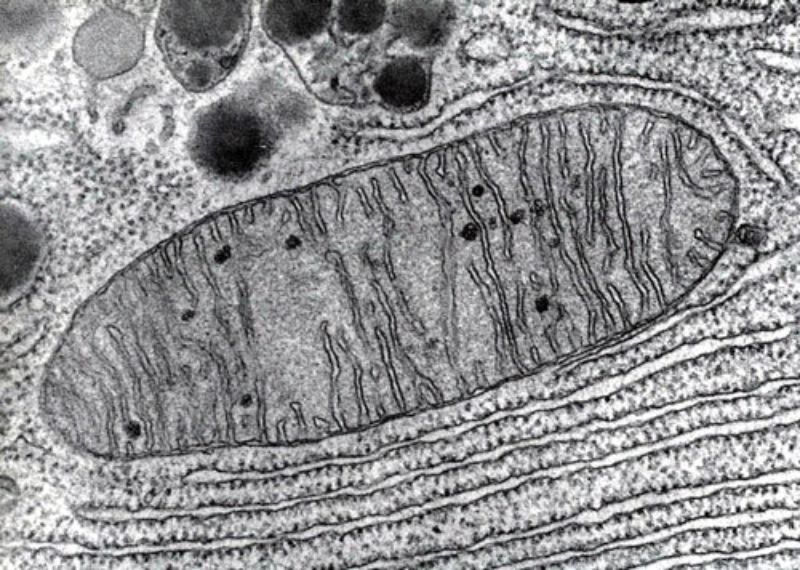
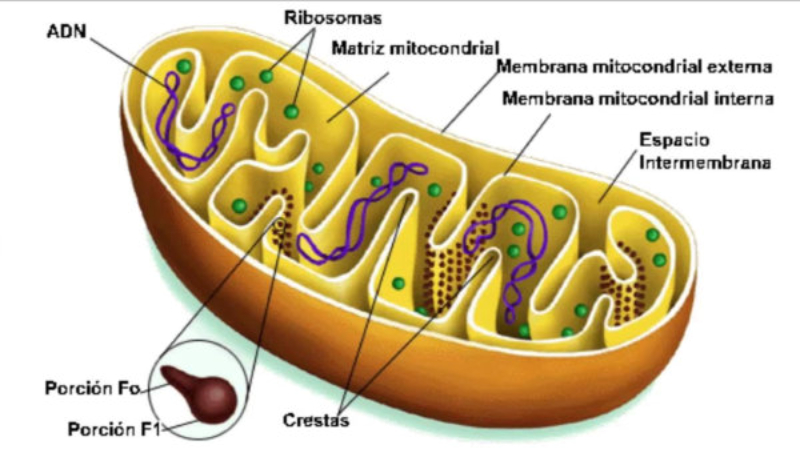
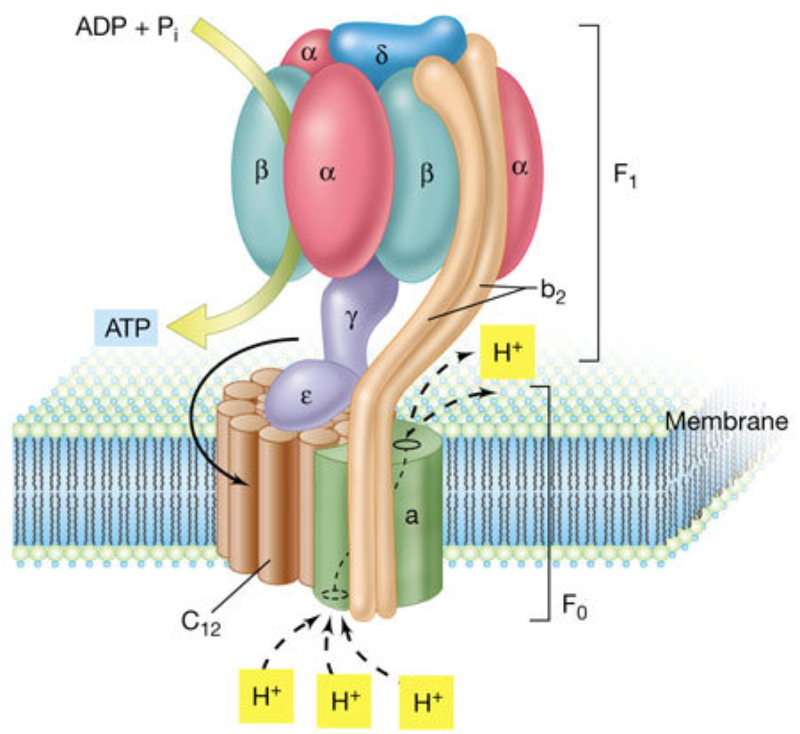
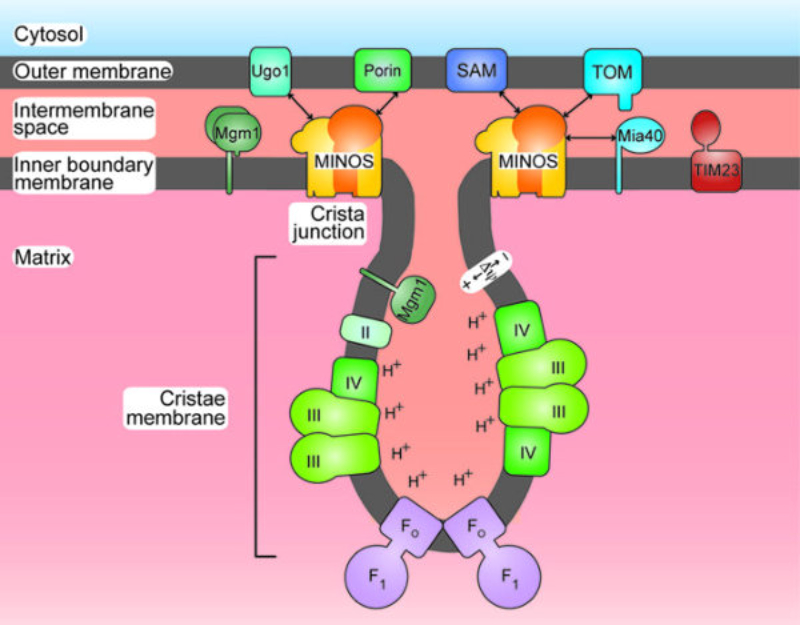
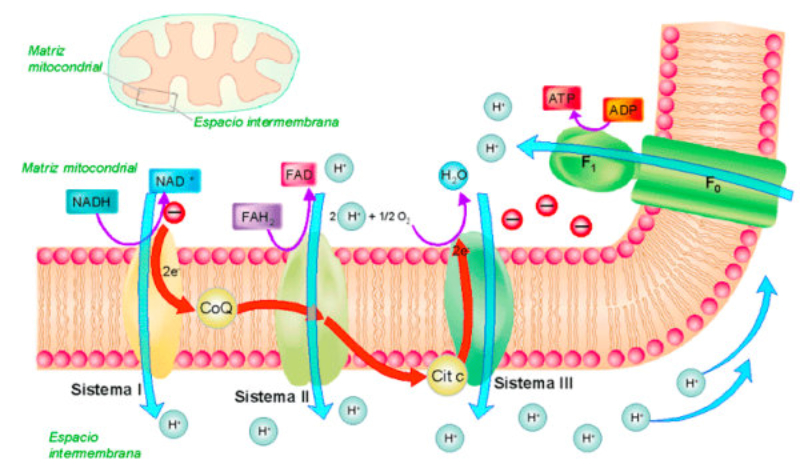
Orgánulos membranosos típicos de eucariotas aerobios, las mitocondrias son las encargadas de realizar la respiración celular. Suelen tener forma alargada, con 1 a 4 µm de longitud. Las mitocondrias tienen dos membranas de diferente estructura y composición: Membrana externa: es lisa y presenta porinas (proteínas de canal para moléculas de bajo peso molecular) y enzimas para diversos procesos metabólicos. Membrana interna: presenta numerosos repliegues llamados crestas mitocondriales (pueden ser laminares, tubulares, prismáticas…). En su composición hay más proteínas que en cualquier otra membrana celular. Además, contiene mucha cardiolipina, un fosfolípido que la hace impermeable a partículas con carga. Inmersa en esta membrana se halla la porción F 0 de la ATP sintetasa, cuya porción F 1 se proyecta hacia la matriz. La presencia de una doble membrana delimita dos espacios en la mitocondria: Espacio intermembranoso: similar al citosol. Matriz mitocondrial: por dentro de la membrana interna. Contienen numerosas proteínas, entre ellas, las del metabolismo oxidativo. También presenta ribosomas de tipo bacteriano, moléculas de ARN y varias cadenas de ADN circular, con información para la síntesis de algunas proteínas mitocondriales. La respiración en las distintas partes de la mitocondria Las mitocondrias son las responsables de la respiración aerobia, proceso en el que se oxida materia orgánica con oxígeno molecular (O 2 ) y se obtiene dióxido de carbono (CO 2 ), agua y energía en forma de ATP. Cada parte de la mitocondria participa de forma diferente en este proceso: Membrana externa: además de las porinas, tiene enzimas para la síntesis de algunos lípidos y la unión de ácidos grasos al coenzima A para que entre en la mitocondria. Espacio intermembranoso: enzimas que fosforilan otros nucleótidos a partir del ATP. Membrana interna: se encarga de la obtención del ATP mediante dos procesos: Cadena respiratoria: formada por enzimas cuyas coenzimas recogen los electrones liberados de la oxidación de la materia orgánica y los llevan al aceptor final, el O 2 . En este proceso se libera energía que se aprovecha para introducir H + en el espacio intermembranoso. Fosforilación oxidativa: aprovecha el gradiente de protones creado para, gracias a la ATP sintetasa, obtener ATP a partir de ADP y fosfato inorgánico (H 3 PO 4 ). Matriz: en ella se da la oxidación de moléculas orgánicas (piruvato, ácidos grasos, aminoácidos) a ácido acético. Además, se realiza el ciclo de Krebs, en el que el acético (en forma de acetil CoA) se oxida a CO 2 .8.6 ESTRUCTURAS EXCLUSIVAS DE CÉLULAS
VEGETALES
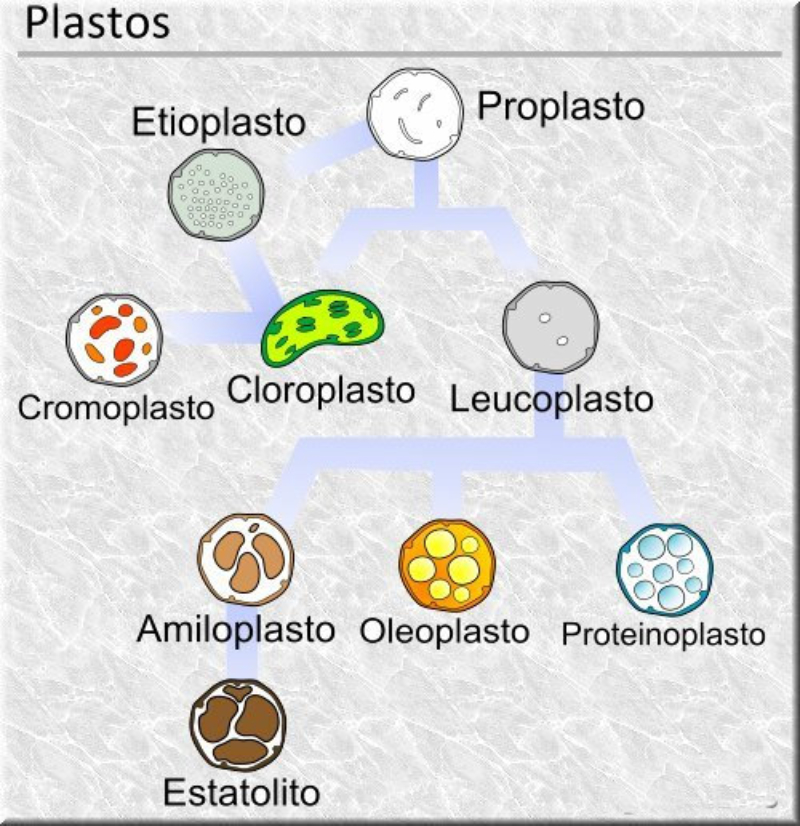
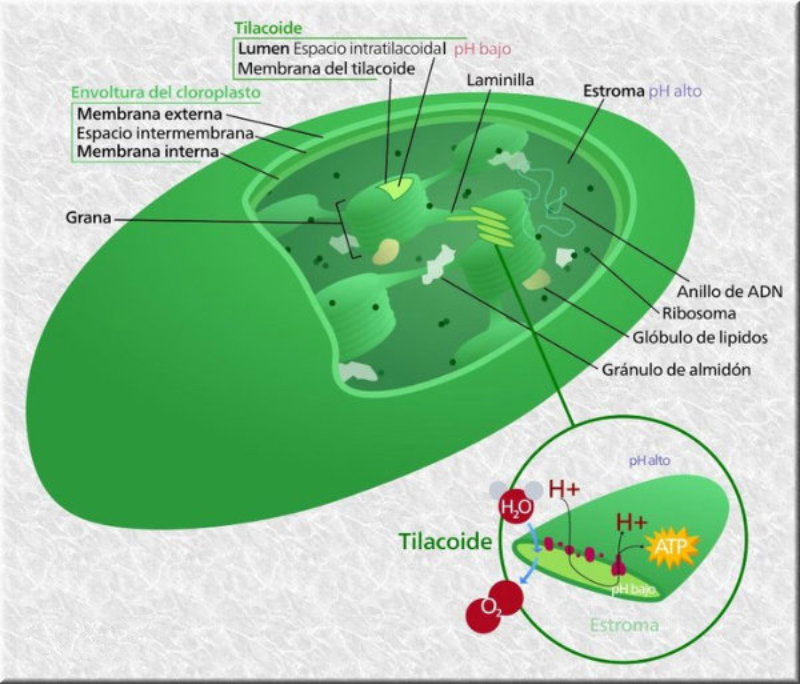
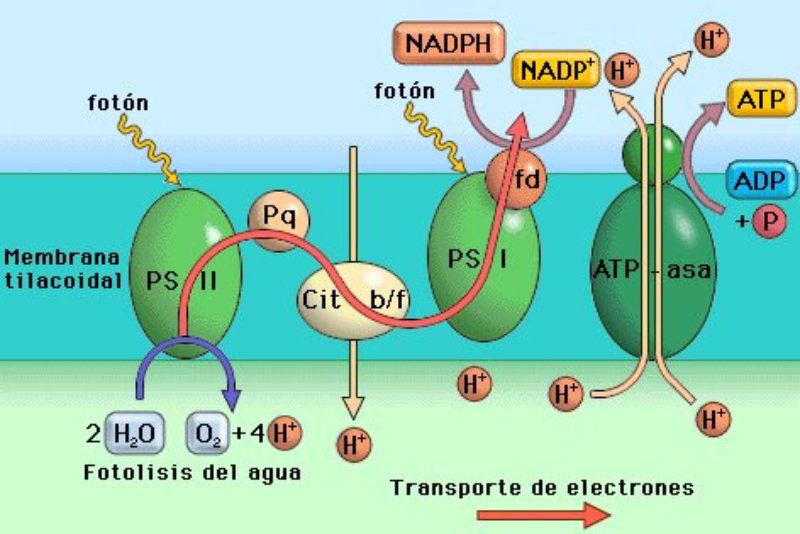
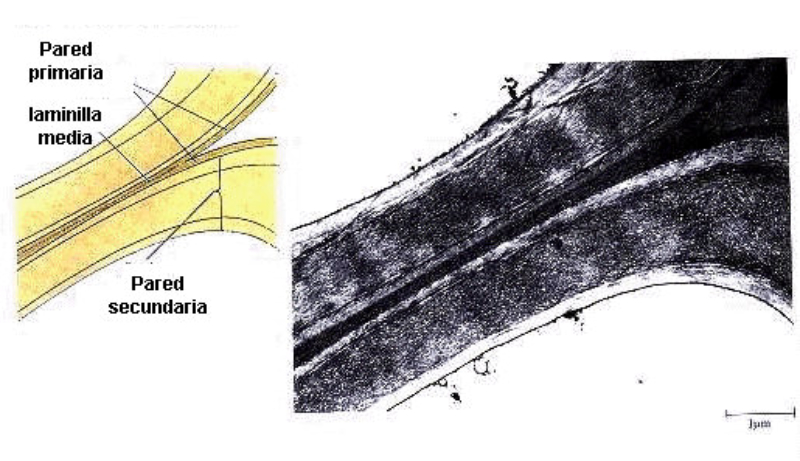
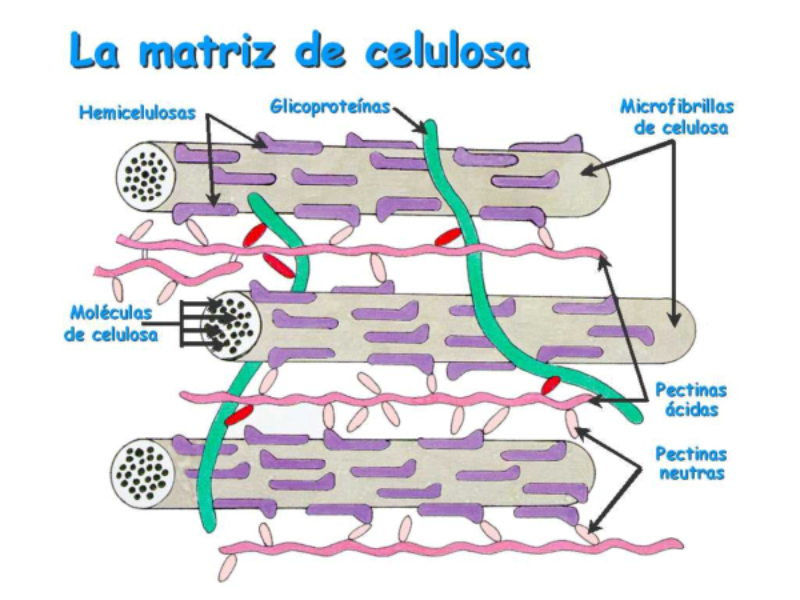
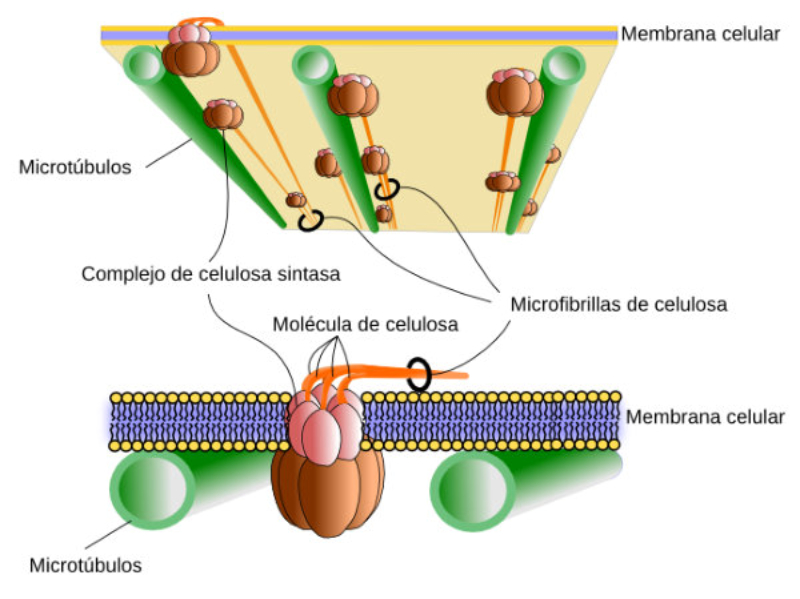
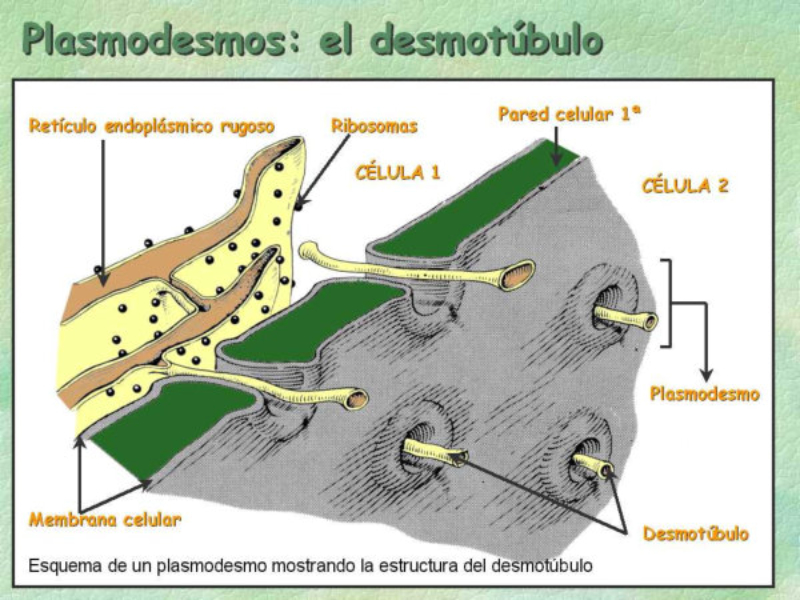
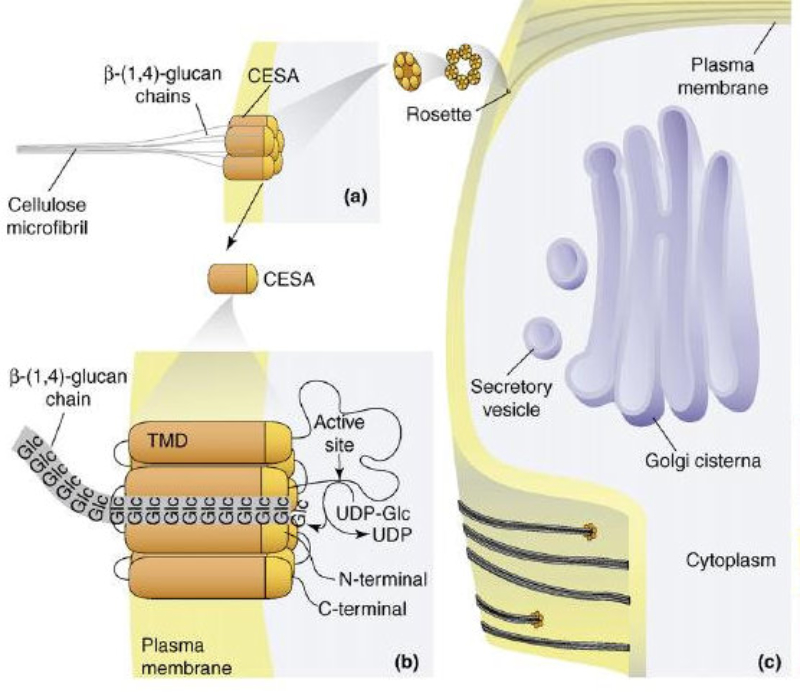
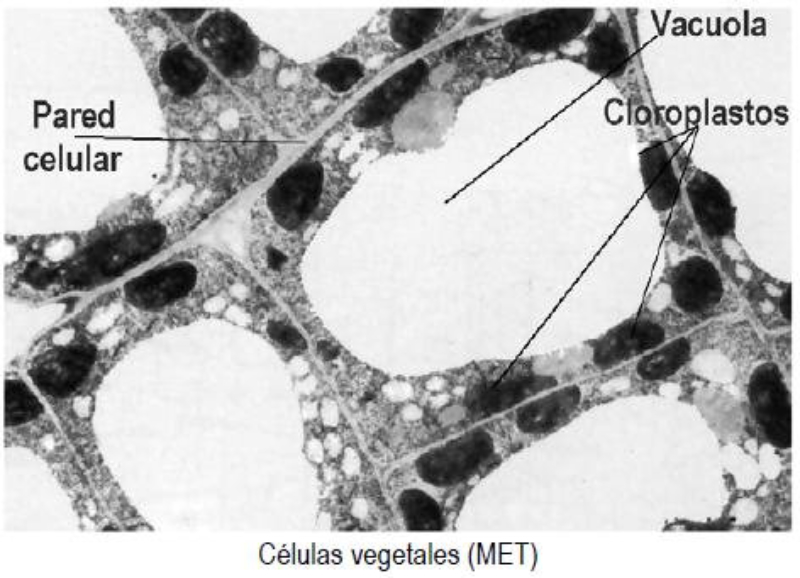
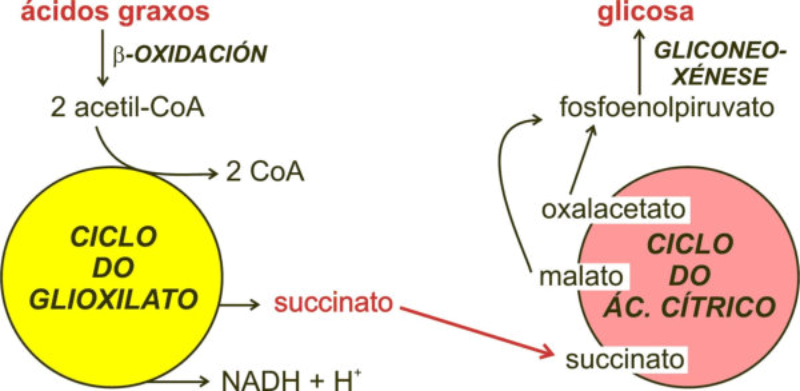
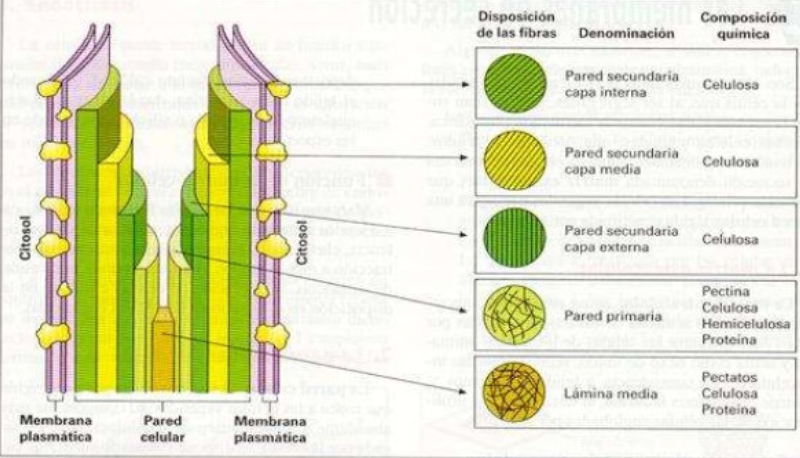
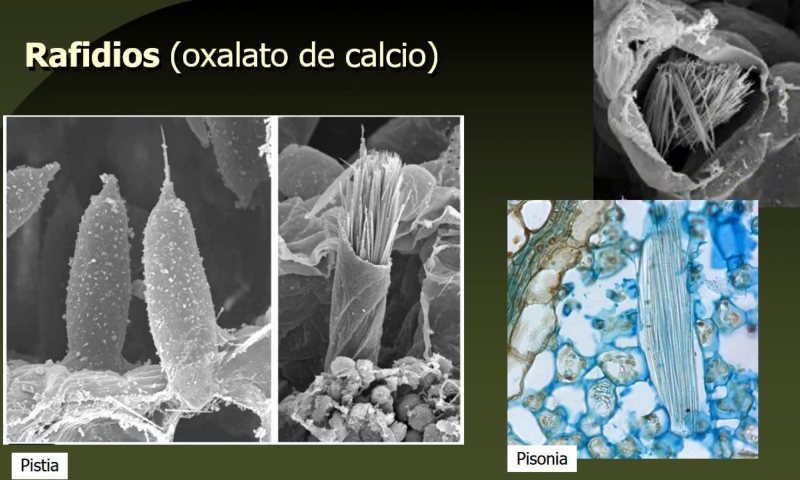
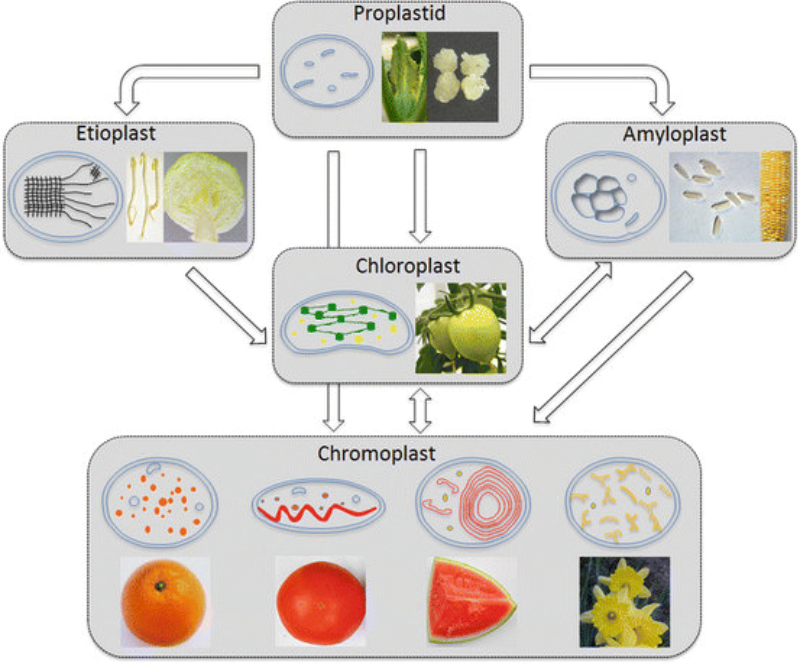
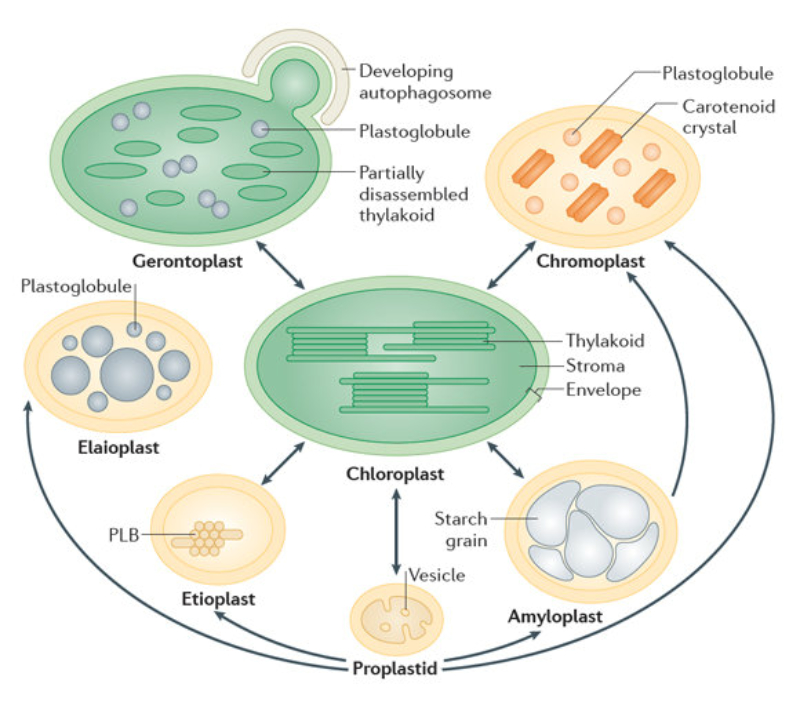
Las células vegetales, además de tener todos los orgánulos de la célula animal (salvo los centríolos), poseen estructuras exclusivas, como plastos, pared celular y vacuolas. Además, en los peroxisomas vegetales se realizan reacciones también exclusivas. 8.6.1. PLASTOS Los plastos, plástidos o plastidios son orgánulos típicos de plantas y algas. Se encargan de producir y almacenar compuestos químicos necesarios para la célula. Básicamente, existen tres tipos de plastos: Leucoplastos: carecen de pigmentos y se hallan en células no expuestas a la luz (raíces, semillas, tubérculos). Se encargan de almacenar almidón (amiloplastos), lípidos (oleoplastos), proteínas (proteoplastos),… Cromoplastos: sintetizan y almacenan pigmentos como carotenos y xantofilas, dando color a frutos, hortalizas, flores… Cloroplastos: contienen numerosos pigmentos, principalmente clorofila, y se encargan de la fotosíntesis. CLOROPLASTOS Los cloroplastos son orgánulos membranosos, generalmente alargados, de 5 a 10 µm de longitud. Tienen tres sistemas de membranas: Membrana externa: muy permeable por la presencia de porinas. Membrana interna: con permeabilidad selectiva gracias a proteínas transportadoras. Carece de pliegues, crestas o cadena transportadora de electrones. Membrana tilacoidal: en el interior del cloroplasto. Forma numerosos sacos aplanados e interconectados, en forma de disco, llamados tilacoides. En ciertas zonas forman pilas de discos llamadas grana. Los tres tipos de membranas determinan tres compartimentos en el cloroplasto: Espacio intermembranoso: entre las membranas externa e interna. Contenido similar al citosol. Estroma: espacio interno del cloroplasto. Contienen numerosas proteínas. También presenta ribosomas de tipo bacteriano, moléculas de ARN, gránulos de almidón, gotas lipídicas y varias cadenas de ADN circular, con información para la síntesis de algunas proteínas del cloroplasto. Espacio tilacoidal: espacio interno de los tilacoides. La fotosíntesis en las diferentes partes del cloroplasto Los cloroplastos se encargan de realizar la fotosíntesis, proceso anabólico que permite la obtención de ATP y moléculas reductoras (NADPH) a partir de la energía solar, con lo que más tarde fabrican moléculas orgánicas. Cada parte del cloroplasto realiza un proceso diferente de la fotosíntesis: Membranas externa e interna: controlan la entrada y salida de sustancias necesarias. Membrana tilacoidal: realizan las reacciones fotosintéticas dependientes de la luz. Los pigmentos localizados en esta membrana capta energía solar, usada para la fotólisis del agua, reacción que proporciona oxígeno molecular que es liberado a la atmósfera y electrones. Una cadena transportadora de electrones lleva los e - hasta la coenzima NADP + , que se reduce a NADPH. Como sucedía en las mitocondrias, la energía liberada en el transporte de e - se emplea para introducir H + desde el estroma al espacio tilacoidal, creando un gradiente electroquímico de protones. Cuando los H + regresen al estroma, lo harán a través de proteínas especiales (ATP sintasa) que utilizarán la energía generada para obtener ATP: fotofosforilación o fosforilación fotosintética. Estroma: realiza las reacciones que no dependen de la luz (ciclo de Calvin). Empleando el ATP y el NADPH generados en la fase luminosa, se reducen (se fijan) moléculas sencillas (CO 2 , NO 3 - , SO 4 2- ) y se elaboran moléculas orgánicas como monosacáridos, aminoácidos, ácidos grasos y bases nitrogenadas. 8.6.2. LA PARED CELULAR La pared celular de las células vegetales puede considerarse una matriz extracelular especial, fabricada por la propia célula y formada principalmente por celulosa. En la pared celular pueden hallarse hasta tres capas diferentes, formadas de forma consecutiva: Lámina media: presente en todas las células. Se forma tras la división celular. Es la capa más externa, común a dos células adyacentes. Formada principalmente por pectina y algo de celulosa, así como sales de Ca + y Mg + . Puede lignificarse en células adultas. Pared primaria: presenta 3 o 4 capas de celulosa con hemicelulosa, aunque su componente principal es una matriz de proteínas y pectinas (heteropolisacáridos que tienden a acumular sales insolubles). Permite el crecimiento celular. Es la capa adyacente a la membrana plasmática si no hay pared secundaria. Pared secundaria: sólo aparece en ciertas células, cuando se ha detenido el crecimiento celular y se produce la especialización. Si está presente, es la capa adyacente a la membrana plasmática. Contiene mucha más celulosa que las otras capas, dispuesta en capas, y se organiza en fibras paralelas con diferente orientación en cada capa. También puede presentar sustancias lipídicas que le dan impermeabilidad (suberina, cutina) u otras que le dan resistencia (lignina), así como sales minerales. Funciones de la pared celular Son muy numerosas y fundamentales para la célula: Soporte mecánico: proporcionan sostén a toda la planta, como un esqueleto. Turgescencia: protege a la célula de fenómenos osmóticos en medio hipotónico, impidiendo que estalle. Protección: frente a la abrasión y el ataque de insectos y patógenos. Comunicación: permite el intercambio de sustancias entre células, sobre todo a través de plasmodesmos, canales citoplasmáticos que comunican células adyacentes. Crecimiento y diferenciación: orienta el crecimiento celular y participa en la diferenciación en tejidos. Formación de la pared celular La pared celular comienza con la lámina media, fina capa péptica que se forma en la división celular, entre las dos células hijas. El engrosamiento posterior se lleva a cabo añadiendo diversos componentes: Proteínas, hemicelulosas y pectinas: las fabrica el retículo endoplasmático y son segregadas por el aparato de Golgi. Igual sucede con sustancias específicas, como la lignina o la suberina. Celulosa: se sintetiza directamente en la superficie celular, mediante la enzima celulosa sintasa, proteína integral de la membrana plasmática. Utiliza glucosas unidas a UDP para formar moléculas de celulosa, que luego une en forma de microfibrillas. Las microfibrillas crecen en una dirección determinada porque la celulosa sintasa se desplaza dirigida por microtúbulos situados bajo la membrana celular. 8.6.3. VACUOLAS Las vacuolas son orgánulos membranosos que, en los vegetales, acumulan sustancias diversas. Pueden ser numerosas en una misma célula o existir sólo una de gran tamaño, ocupando incluso el 90% del volumen celular. La membrana de las vacuolas se denomina tonoplasto y presenta sistemas de transporte activo para: El bombeo de iones: se introducen iones en el espacio vacuolar, lo que hace que entre agua por ósmosis y ayuda a la turgencia junto con la resistencia de la pared celular. A veces, la concentración de sales es tan alta que precipitan en cristales, como los de oxalato cálcico, que adoptan formas muy diversas. El bombeo de protones (H + ): la introducción de H + hace descender el pH de la vacuola, de forma similar a lo que sucede en los lisosomas. Funciones de las vacuolas Almacén de sustancias: agua, azúcares, aminoácidos, proteínas, polisacáridos, iones, desechos,… Acumulación de pigmentos: sobre todo antocianinas (azul, rojo, púrpura), que dan color a frutas y verduras. Regulación osmótica: por la acumulación de iones y agua. Digestión intracelular: similar a los lisosomas. Digieren ribosomas, mitocondrias, plastidios, macromoléculas… cuyos componentes son después reciclados. Defensa: acumulan sustancias tóxicas (como la nicotina) que luego pueden liberar si la célula es dañada por parásitos o herbívoros. 8.6.4. PEROXISOMAS ESPECIALES Las plantas presentan algunos peroxisomas con funciones muy específicas: Glioxisomas: realizan el ciclo del glioxilato, en el que los lípidos se convierten en azúcares. Son esenciales para la germinación de semillas de oleaginosas. Fotorrespiración: en las hojas, los peroxisomas se asocian a mitocondrias y cloroplastos y realizan un proceso relacionado con la fotosíntesis, la fotorrespiración.




























Para ir a donde no se sabe hay que ir por donde no se sabe.” San Juan de la Cruz
“It must be a strange world not being a scientist, going through life not knowing--or
maybe not caring about where the air came from, where the stars at night came from or
how far they are from us. I WANT TO KNOW” Michio Kaku
“Nullius in verba” Robert Boyle, Christopher Wren y Robert Hooke







Ramón y Cajal y Camillo Golgi
Premios Nobel de Fisiología en 1906.





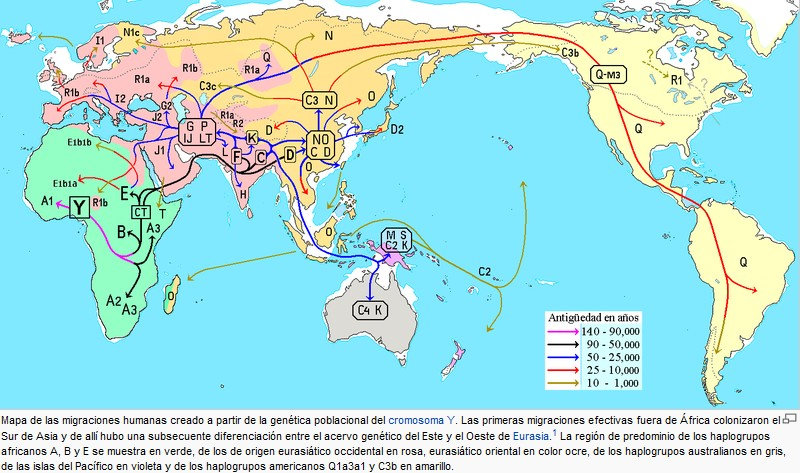
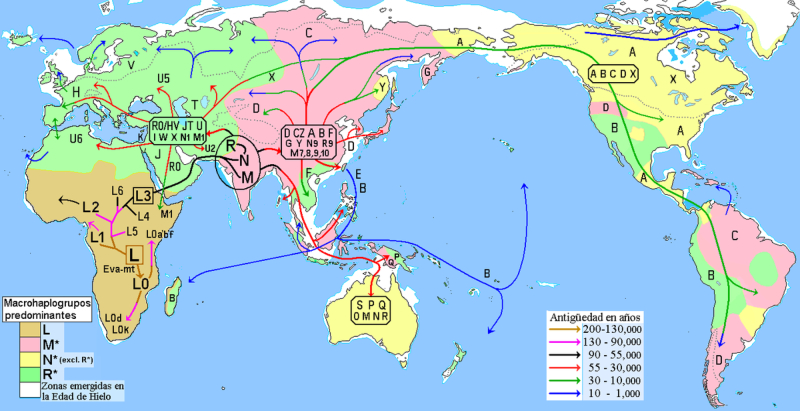
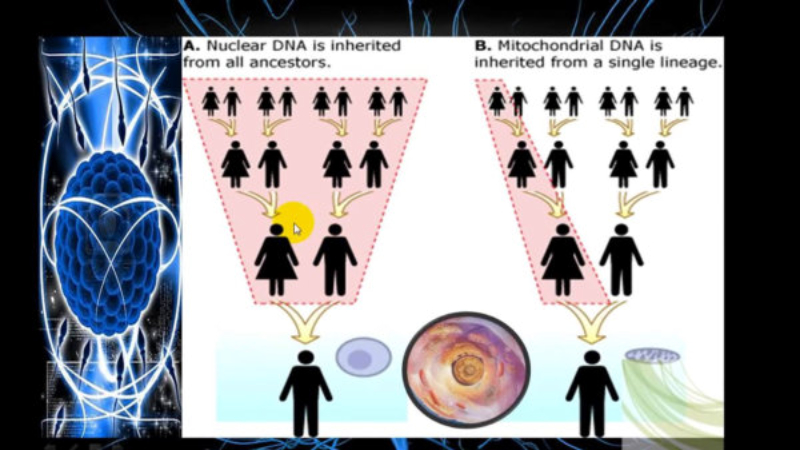
Mapa de la hipótesis de las migraciones humanas, basada en el origen y
dispersión de los haplogrupos del ADN mitocondrial.































Paisajes celulares
(Interactivo)








LA CÉLULA: MORFOLOGÍA CELULAR

![[x]](bg2bto_htm_files/close.png "Close")