BIOLOGÍA Y GEOLOGÍA
METABOLISMO
5. PROCESOS ANABÓLICOS
La célula realiza multitud de procesos anabólicos, entre los que destacan: Biosíntesis de aminoácidos y proteínas: algunos aa pueden ser sintetizados a partir de precursores. Los aa esenciales deben tomarse con la dieta. A partir de estos aa se sintetizan las proteínas: traducción en ribosomas. Biosíntesis de ácidos nucleicos: ADN y ARN. Duplicación y transcripción. Biosíntesis de ácidos grasos o lipogénesis: Se realiza en el citosol, a partir de acetil-CoA procedente del catabolismo de otras biomoléculas. También se sintetizan otros lípidos. Biosíntesis de glucógeno: glucogenosíntesis. En el citosol, a partir de glucosa-6-fosfato. Gluconeogénesis: síntesis de glucosa a partir de moléculas más sencillas. Las plantas pueden sintetizar azúcares a partir de ácidos grasos mediante el ciclo del glioxilato, en los glioxisomas.5.1 GLUCONEOGÉNESIS
Básicamente es un proceso inverso a la glucólisis, salvo algunas reacciones que requieren rutas diferentes y gran aporte de energía. Se parte de 2 moléculas de pirúvico. El pirúvico debe convertirse en 2- fosfoenol-pirúvico. Esta es la etapa más compleja, pues requiere varias reacciones, energía en forma de ATP y GTP y poder reductor como NADH + H + . A partir del fosfoenol-pirúvico las reacciones son las mismas que en la glucólisis y con casi las mismas enzimas, pero en sentido inverso.6. ANABOLISMO AUTÓTROFO: FOTOSÍNTESIS
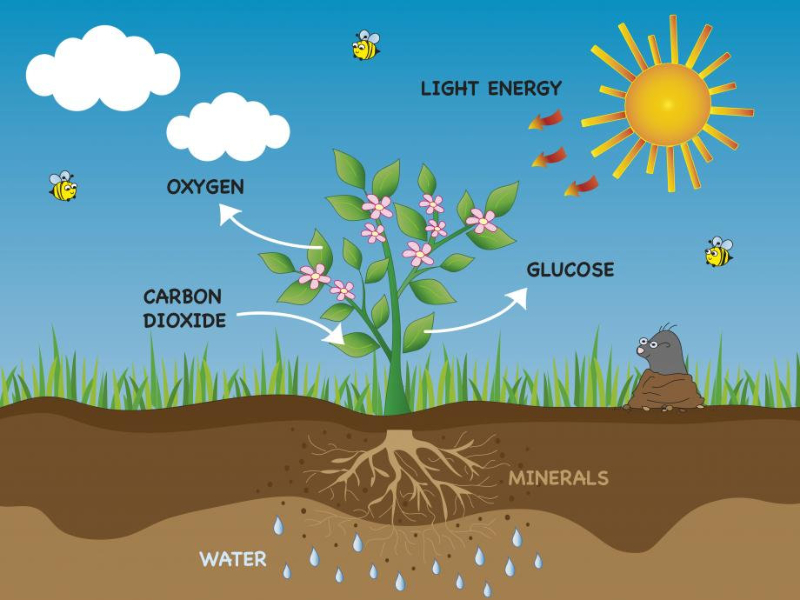
La fotosíntesis es un proceso anabólico para obtener materia orgánica a partir de la inorgánica y con ayuda de la energía lumínica. Se realiza en cloroplastos de células eucariotas de organismos fotosintéticos. La fotosíntesis consta de 2 fases denominadas fase lumínica y fase oscura.6.1. FASE LUMÍNICA O FOTOQUÍMICA
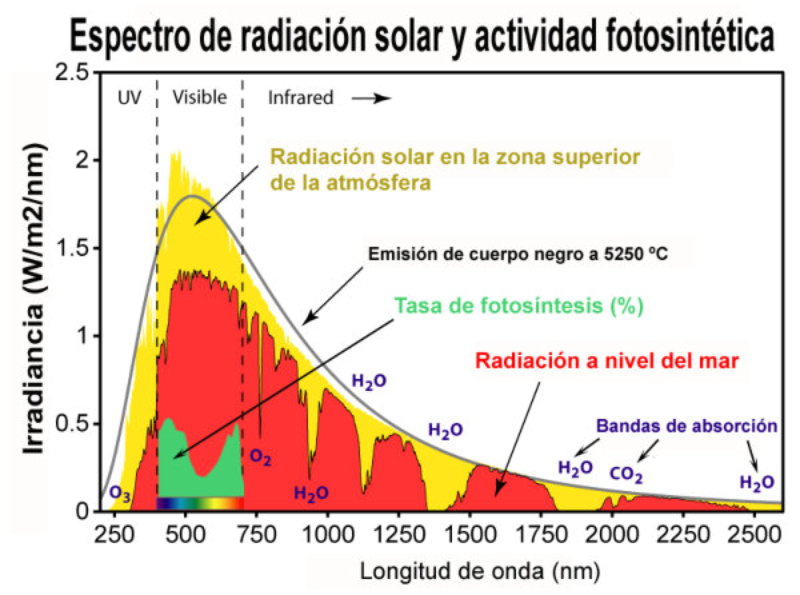
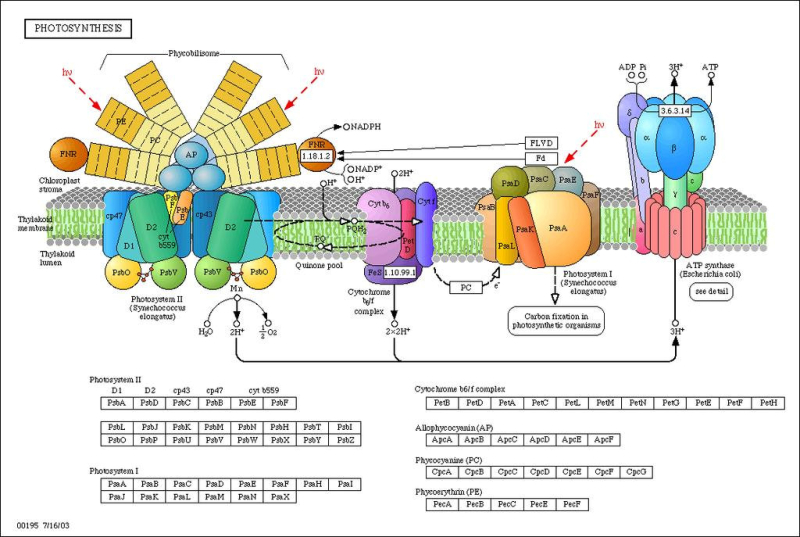
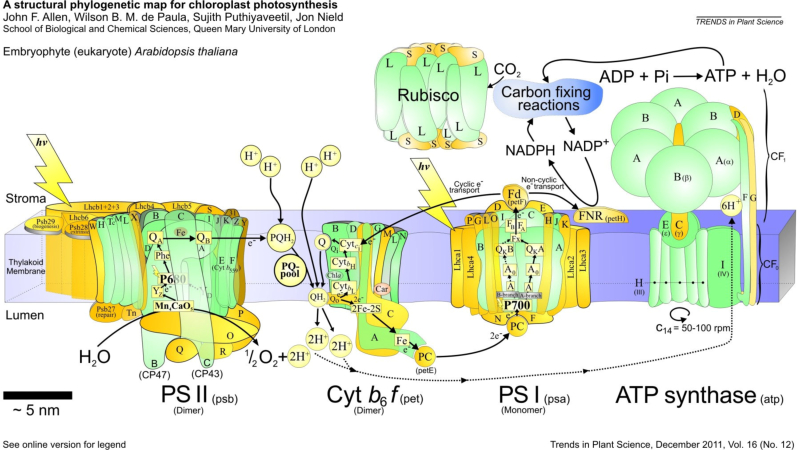
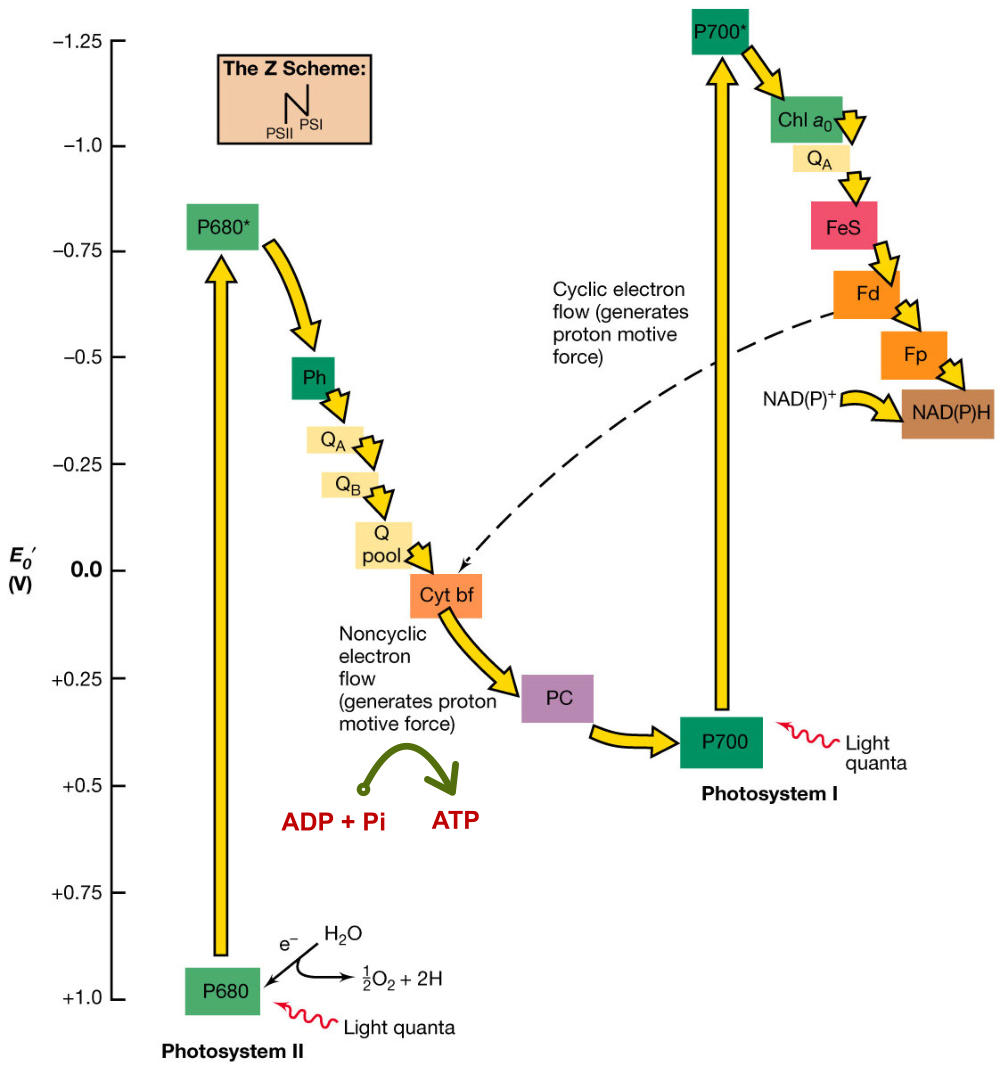
La fase lumínica o fotoquímica consiste en una serie de reacciones fotoquímicas en las membranas de los tilacoides. Se capta energía lumínica y se transforma en energía química. La incidencia de fotones sobre los fotosistemas libera electrones que reducen el NADP + a NADPH (poder reductor) y permiten la síntesis de ATP (fotofosforilación). Los electrones cedidos son aportados por un dador de electrones, que en las plantas es el H 2 O, liberando O 2 (fotosíntesis oxigénica). FOTOSISTEMAS Son la unidad estructural de la membrana tilacoidal. Están formados por: Un complejo antena: son cientos de moléculas de clorofila y otros pigmentos, unidos a proteínas de la membrana, que captan luz de diferente longitud de onda y transfieren su energía al centro reactivo. Centro reactivo o centro de reacción fotoquímica: formado por dos moléculas especiales de clorofila dentro de una proteína transmembrana. Actúa como una trampa energética, captando la energía que llega del complejo antena y usándola para impulsar electrones hacia la cadena transportadora de electrones de la membrana tilacoidal. Un dador y un aceptor de electrones: diferentes en cada fotosistema. TIPOS DE FOTOSISTEMAS Los vegetales superiores presentan en sus membranas tilacoidales dos tipos de fotosistemas: fotosistema I (PS I) y fotosistema II (PS II). Ambos están conectados por una cadena transportadora de electrones. PS I: se halla por toda la membrana tilacoidal. Contiene dos moléculas de clorofila P 700 (absorbe el máximo de luz a 700 nm). La clorofila P 700 cede sus electrones a un aceptor primario que es una molécula llamada clorofila A 0 . El hueco electrónico se llena con un electrón procedente del dador, la plastocianina (PC), que se encuentra en la cadena transportadora de electrones. PS II: se encuentra principalmente en los grana. Contiene dos moléculas de clorofila P 680 , las cuales ceden su electrón a un aceptor primario que es la feofitina (Feo). El hueco se rellena con un electrón procedente del dador, el agua. Por tanto, en este fotosistema sucede la fotolisis del agua, en la que una molécula de agua se rompe dando dos electrones, dos H + y O 2 que se desprende. En los fotosistemas, los complejos antena actúan como auténticas antenas recolectoras de fotones, que producen electrones energéticos que son transportados hasta el centro reactivo, el cual libera un electrón de alta energía. PIGMENTOS FOTOSINTÉTICOS Todos los organismos fotosintéticos tienen varios tipos de pigmentos para captar la energía lumínica. Entre ellos: Clorofilas: contienen una porfirina unida a un átomo de Mg 2+ y a una cadena de fitol, lípido isoprenoide que permite su unión con fosfolípidos y regiones hidrófobas de las proteínas de las membranas tilacoidales. Existen numerosas variedades, pero las principales son la clorofila a y la clorofila b, que absorben luz roja y azul. Carotenoides (carotenos y xantofilas): absorben luz verde y azul, protegiendo a las clorofilas de procesos de oxidación. Ficobilinas: en ciertas algas y bacterias. Ficocianina, ficoeritrina,... Absorben luz verde y amarilla (ficobilisomas). FOTOFOSFORILACIÓN NO CÍCLICA (OXIGÉNICA) En esta fotofosforilación los dos PS actúan en serie y los e - siguen un flujo lineal desde el agua hasta el NADP + , que se reduce a NADPH. El flujo electrónico genera energía con la que se produce ATP. La representación de este proceso se denomina esquema Z: Los fotones que llegan al PS II son recolectados por su complejo antena y se llevan al centro P 680 , que libera un e - de alta energía. Este e - pasa por la cadena de transporte (feofitina, plastoquinona, cit-b 6 -f y plastocianina). Los e - llegan finalmente al PS I. Durante este transporte se libera energía que se emplea en producir ATP a partir de ADP + P i : fotofosforilación. Los huecos electrónicos dejados en el PS II son rellenados por e - del H 2 O (dador de e - ), que sufre fotólisis y libera O 2 , e - y protones. Los fotones que llegan al PS I son captados por el P 700 , que libera un e - de alta energía, que será transportado por otra cadena (clorofila A 0 , filoquinona (Q), ferredoxina y ferredoxina-NADP- reductasa). Finalmente, acaba en el NADP que se reduce a NADPH. Los huecos electrónicos del PS I son rellenados por los e - que llegan del PS II (la plastocianina es, pues, el dador de e - del PSI. HIPÓTESIS QUIMIOSMÓTICA DE LA FOTOFOSFORILACIÓN Esta hipótesis explica la formación de ATP de forma similar a como sucede en las mitocondrias: La fotólisis del agua en el PS II se produce en el estroma. Dos de los e - y 2 de los H + liberados son aceptados por la plastoquinona (PQ), que se reduce a PQH 2 . La PQH 2 pasa los e - al cit-b 6 -f y los H + al espacio tilacoidal. Por tanto, el complejo plastoquinona y cit-b 6 -f actúa como una bomba de protones hacia el espacio tilacoidal, generando un gradiente electroquímico. Según la hipótesis quimiosmótica de Mitchell, el flujo de protones desde el espacio tilacoidal hasta el estroma tiene lugar a través de la enzima ATP sintetasa, que genera ATP por catálisis rotacional. FOTOFOSFORILACIÓN CÍCLICA (ANOXIGÉNICA) En esta fotofosforilación sólo interviene el PS I y los e - del P 700 realizan un recorrido cíclico y vuelven a este mismo centro reactivo. Lo e - pasan a la A 0 , filoquinona, ferredoxina, plastoquinona y cit-b 6 -f. Este último libera suficiente energía para crear un gradiente de protones que permite la síntesis de ATP por la ATP sintetasa. Finalmente, los e - del cit-b 6 -f pasan a la plastocianina, que los devuelve al PS I. Por tanto, en este proceso se obtiene ATP, pero no O 2 ni NADPH, logrando un suplemento extra de ATP.6.2. FASE OSCURA O BIOSINTÉTICA
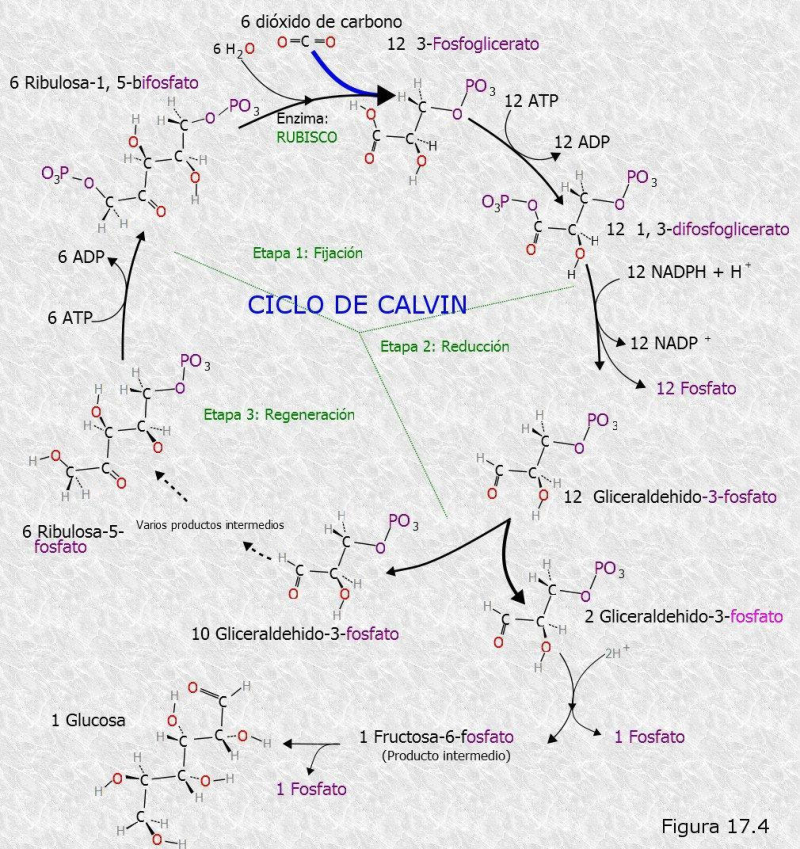
Esta fase transcurre en el estroma del cloroplasto y es independiente de la luz. Se emplea el ATP y el poder reductor (NADPH) obtenidos en la fase luminosa para reducir moléculas sencillas oxidadas (CO 2 , NO 3 - , SO 4 2- ,...). Así se consiguen moléculas orgánicas sencillas que se utilizarán en otros procesos. EL CICLO DE CALVIN-BENSON Es un proceso de fijación y reducción del CO 2 atmosférico. Es un proceso anabólico y reductor y, por tanto, precisa energía (ATP) y poder reductor (NADPH). El proceso comienza cuando el CO 2 es fijado y unido a una molécula de 5C, la ribulosa-1,5-bifosfato (RuDP), originando una molécula de 6C muy inestable que se escinde rápidamente en 2 moléculas de ácido 3- fosfoglicérico (3PG). Esta reacción está catalizada por la enzima ribulosa-1,5 difosfato carboxilasa oxigenasa (Rubisco). Esta es una enzima “lenta” (cataliza unas 3 moléculas de sustrato por segundo, frente a las 1000 típicas de otras enzimas). Por ello se necesita en grandes cantidades. De hecho, alrededor del 50% de las proteínas del cloroplasto son rubisco, lo que la convierte en la proteína más abundante de la naturaleza. El ciclo de Calvin y la fijación del CO2 por la rubisco son propios de todos los organismos autótrofos. No se dan sólo en fotosintetizadores típicos, como cianobacterias, algas eucarióticas y plantas, que realizan la fotosíntesis oxigénica, sino también en arqueas y bacterias dotadas de metabolismos fotosintetizadores o quimiosintetizadores diversos. Seguidamente, el 3PG se reduce a gliceraldehído-3-fosfato (G-3-P) mediante ATP y NADPH. El G-3-P puede ahora seguir dos rutas: 10 de cada 12 moléculas de G-3-P seguirán el ciclo de Calvin y, tras varias reacciones en las que intervienen triosas, tetrosas, pentosas, hexosas y heptosas, se regenera de nuevo la RuDP. 2 moléculas de G-3-P pasan al citosol, donde son precursoras de la síntesis de glucosa (gluconeogénesis), con la que se formará sacarosa, almidón, ácidos grasos y aminoácidos. La síntesis de una molécula de glucosa precisa de 6 CO 2 , 6 moléculas de H 2 O, 18 ATP y 12 NADPH. 6 CO 2 + 6 H 2 O + 18 ATP + 12 NADPH C 6 H 12 O 6 (glucosa) + 6 O 26.3. FOTORRESPIRACIÓN Y PLANTAS C
4
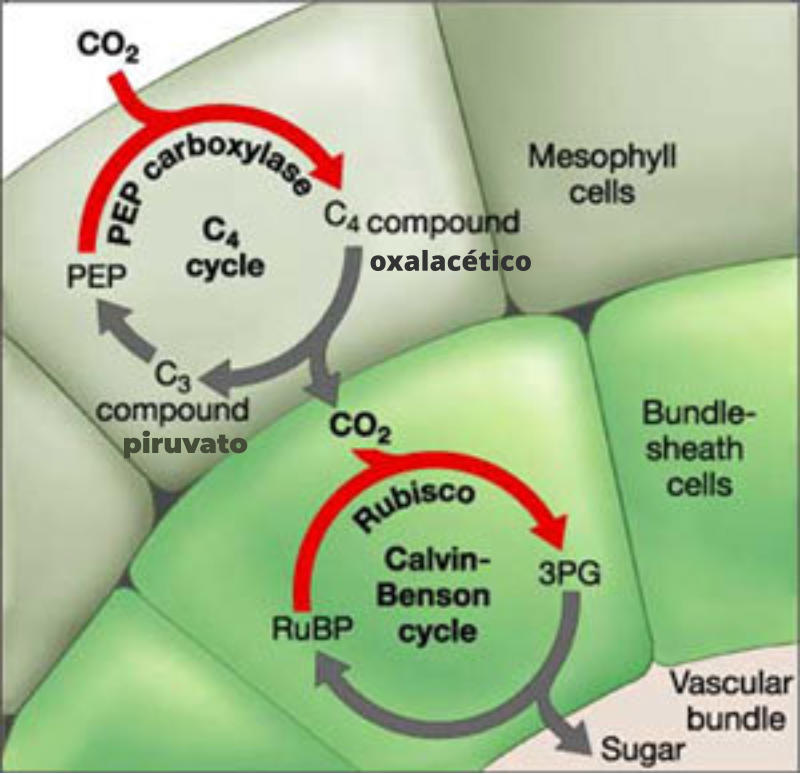
La rubisco es una enzima poco eficaz. No sólo por su lentitud catalítica, sino también porque, además de carboxilasa es oxidasa. Por ello, puede unir CO 2 a la RuDP para el ciclo de Calvin-Benson, pero también puede añadirle O 2 y dar fotorrespiración El que realice una u otra reacción depende de la concentración relativa de CO 2 y O 2 . Si [CO 2 ]>[O 2 ], la rubisco actúa como carboxilasa: fija una molécula de CO 2 a la RuDP y forma dos moléculas de 3PG. Si [CO 2 ]<[O 2 ], la rubisco actúa como oxigenasa: incorpora O 2 a la RuDP, que se rompe en una molécula de 3GP (que pasa al ciclo de Calvin) y otra de ácido fosfoglicólico, que pasa a los glioxisomas y a las mitocondrias, liberando una parte de sus átomos de carbono en forma de CO 2 . Esto supone una pérdida de la eficacia del proceso fotosintético. Como en este proceso se consume O 2 , se libera CO 2 y sólo sucede en presencia de luz, se denomina fotorrespiración. No se conoce bien para qué sirve la fotorrespiración, pero parece ser que, en parte, protege a los tilacoides de la fotooxidación que sufren ante un exceso de luz y escasez de CO 2 . PLANTAS C 4 : SOLUCIÓN A LA FOTORRESPIRACIÓN Las plantas que fijan directamente el CO 2 a la RuDP y forman 3PG se denominan plantas C 3 , por ser de 3 carbonos estas primeras moléculas. Son plantas con gran fotorrespiración. Sin embargo, existen plantas que fijan primero el CO 2 a una molécula de 3 carbonos (el ácido fosfoenolpirúvico: PEP), dando una molécula de 4 carbonos (el ácido oxalacético). Estas plantas se llaman C 4 e incluyen a las plantas adaptadas a climas secos y cálidos: cañas, maíz, mijo, papiro y plantas CAM (Crassulacean Acid Metabolism) (cactáceas y plantas crasas). En estas plantas tiene lugar la fijación del CO 2 en las células del mesófilo de la hoja. Allí se da el ciclo de Hatch-Slack: el CO 2 se une al PEP (3C) y forma oxalacético (4C). La enzima que cataliza esta reacción es mucho más activa que la rubisco. El oxalacético pasa luego a las células que rodean a los vasos conductores de la hoja, en cuyos cloroplastos tiene lugar la fotosíntesis. Allí el oxalacético se descarboxila y libera PEP (que vuelve al mesófilo) y CO 2 (que va al ciclo de Calvin). De esta forma se incrementa la concentración de CO 2 y se evitan las pérdidas por fotorrespiración.6.4. FACTORES QUE MODULAN LA FOTOSÍNTESIS
El rendimiento de la fotosíntesis depende de numerosos factores ambientales, entre ellos: Intensidad lumínica: cada especie tiene un intervalo de luminosidad óptimo. Dentro de ese intervalo, el aumento de la luminosidad aumenta la fotosíntesis hasta un máximo. [CO 2 ]: a medida que aumenta, aumenta la actividad fotosintética, hasta llegar a un límite estable. [O 2 ]: su incremento disminuye la fotosíntesis debido a la fotorrespiración, sobre todo en plantas C 3 . Fotoperiodo: el rendimiento fotosintético depende de las estaciones, que proveen un número de horas de luz muy variable. Temperatura: en general, el aumento de la temperatura incrementa la fotosíntesis, pues aumenta la actividad enzimática. Por encima de cierto valor, la fotosíntesis disminuye. De todas formas, hay plantas adaptadas a climas cálidos y otras a climas fríos. Humedad: la falta de humedad provoca el cierre de los estomas y la disminución de la actividad fotosintética. Las plantas C 4 son más eficaces en condiciones secas. Color de la luz: por encima de una longitud de 680 nm (luz roja) el PS II no actúa, por lo que sólo se da fotofosforilación cíclica.7. ANABOLISMO AUTÓTROFO:
QUIMIOSÍNTESIS
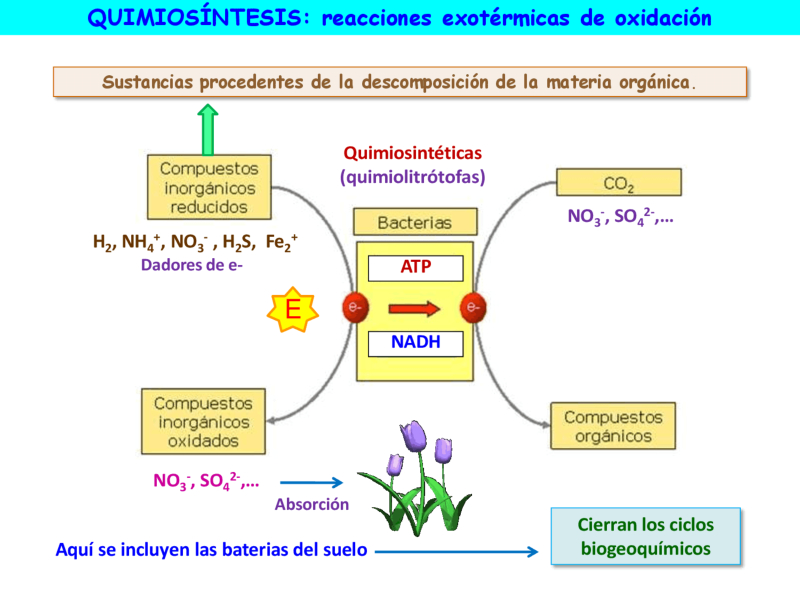
La quimiosíntesis consiste en la síntesis de ATP a partir de la energía que se libera en reacciones de compuestos inorgánicos reducidos. Los organismos que realizan quimiosíntesis se denominan quimoautótrofos, quimiolitótrofos o quimiosintéticos, todos son bacterias que usan como fuente de carbono el dióxido de carbono en un proceso parecido al ciclo de Calvin de las plantas. La ecuación global de la quimiosíntesis, usando como molécula oxidable el sulfuro de hidrógeno es la siguiente: CO 2 + O 2 + 4 H 2 S CH 2 O (formaldehído) + 4 S + 3 H 2 O Muchas bacterias que viven en el fondo de los océanos usan la quimiosíntesis como forma de producir energía sin necesidad de luz solar, donde no pueden vivir organismos fotosintéticos. Muchas de estas bacterias son la fuente básica de alimentación para el resto de organismos del suelo oceánico, siendo el comportamiento simbiótico muy común. Muchos de los compuestos reducidos que utilizan las bacterias, como el NH 3 o el H 2 S son sustancias procedentes de la descomposición de la materia orgánica. Al oxidarlas, las transforman en sustancias minerales, NO 3- y SO 4 2- , respectivamente, que pueden ser absorbidas por las plantas. Estas bacterias cierran, por tanto, los ciclos biogeoquímicos, posibilitando la vida en el planeta.7.1 FASES DE LA QUIMIOSÍNTESIS
La quimiosíntesis consta de dos fases: Fase oxidativa: es similar a la fase fotoquímica o luminosa de la fotosíntesis. En esta fase, la energía liberada en las reacciones de oxidación de sustancias inorgánicas (amoniaco, nitritos, azufre, hierro, etc.) se utiliza para sintetizar ATP (fosforilación oxidativa del ADP) y poder reductor (NADH en las bacterias en lugar de NADPH como en las plantas). Fase biosintética: es similar a la fase biosintética u oscura de la fotosíntesis. Utiliza el ATP y el NADH obtenidos en la fase anterior para reducir compuestos inorgánicos (CO 2 , NO -3 , SO 2 -4 ) y obtener compuestos orgánicos.7.2 TIPOS DE BACTERIAS QUIMIOSINTÉTICAS
Según el sustrato utilizado, las bacterias se clasifican en los siguientes grupos: Bacterias incoloras del azufre: Estas bacterias usan el H 2 S procedente de la descomposición de la materia orgánica, que abunda en las aguas residuales, fuentes hidrotermales y ambientes ricos en azufre o H 2 S. Bacterias del nitrógeno: Viven en el suelo y en el agua. Oxidan compuestos reducidos de nitrógeno, en concreto el amoniaco (NH 3 ), que transforman en nitratos (NO 3- ), el cual puede ser utilizado por las plantas. Esta oxidación del amoniaco procedente de la descomposición de la materia orgánica se realiza en dos fases: Bacterias nitrosificantes: Oxidan el amoniaco a nitritos. Bacterias nitrificantes: Oxidan los nitritos a nitratos. Bacterias del hierro: Oxidan compuestos ferrosos a férricos. Bacterias del hidrógeno: Quimioautótrofas facultativas, que pueden utilizar el hidrógeno molecular.





























Para ir a donde no se sabe hay que ir por donde no se sabe.” San Juan de la Cruz
“It must be a strange world not being a scientist, going through life not knowing--or
maybe not caring about where the air came from, where the stars at night came from or
how far they are from us. I WANT TO KNOW” Michio Kaku
“Nullius in verba” Robert Boyle, Christopher Wren y Robert Hooke








































Fotosistemas con punto de cruz












![[x]](bg2bto_htm_files/close.png "Close")